Clear Sky Science · pt
Tioredoxina e sua proteína parceira são essenciais para a formação do flagelo de zoósporos em Actinoplanes missouriensis
Como algumas bactérias constroem rabos minúsculos sob demanda
Muitas bactérias se movem usando longos apêndices em forma de chicote chamados flagelos. Para um microrganismo do solo chamado Actinoplanes missouriensis, construir esses apêndices no momento exato é uma questão de sobrevivência: seus esporos precisam se tornar nadadores assim que há chuva ou água disponível. Este artigo revela como um par de pequenas proteínas age como um interruptor molecular liga–desliga para controlar quando esses apêndices de natação são formados.
De esporos silenciosos a nadadores repentinos
A. missouriensis normalmente cresce como filamentos ramificados no solo. Em condições secas, ele forma estruturas arredondadas chamadas esporângios nas pontas desses filamentos. Dentro de cada esporângio, centenas de esporos redondos se formam e se preparam para a vida fora dali. Quando a água chega, o esporângio se abre, liberando os esporos no líquido. Esses esporos rapidamente desenvolvem flagelos e se tornam “zoósporos” que podem nadar por um curto período, ajudando-os a se dispersar para novos locais antes de se estabelecerem e crescerem novamente. Como essa fase de natação é breve e rigorosamente cronometrada, a célula deve controlar com precisão quando e como os flagelos são montados.
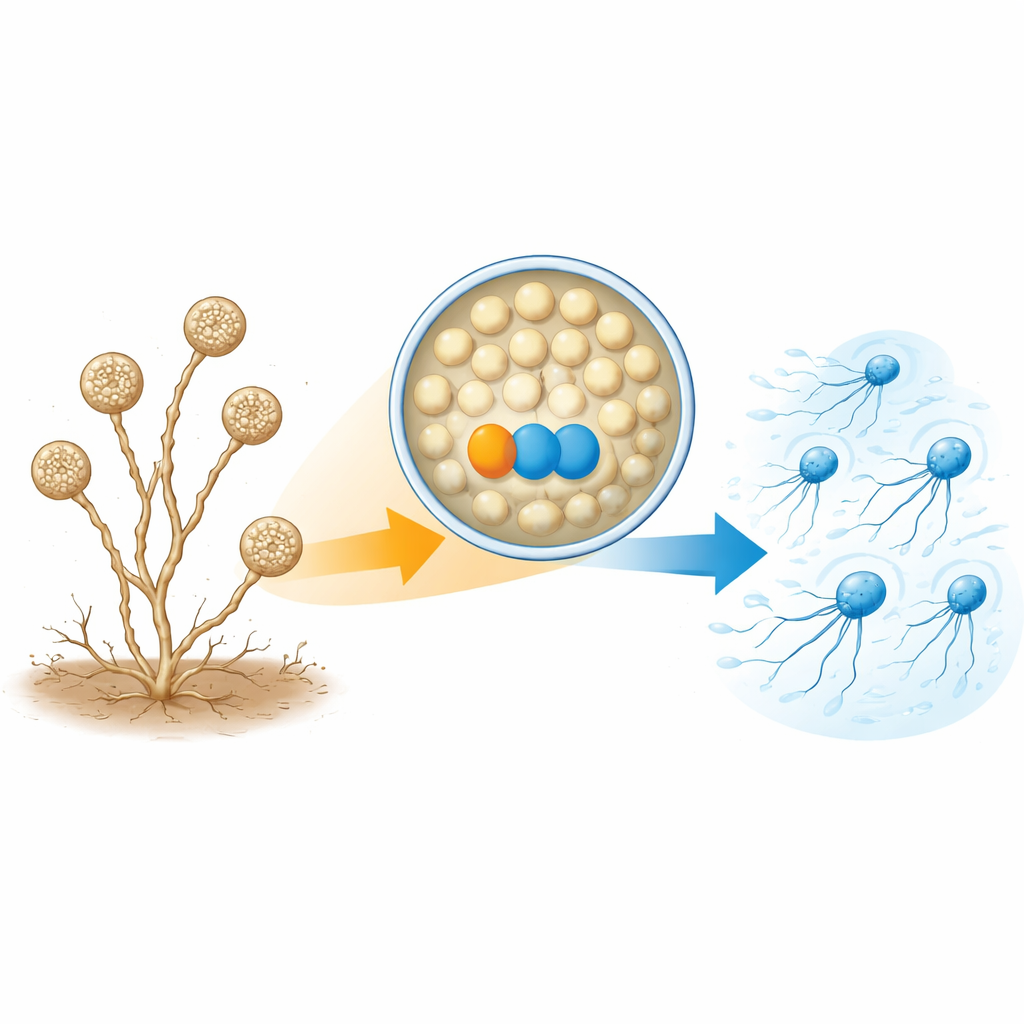
Um par de proteínas necessário para construir os apêndices
Os autores concentraram-se em duas proteínas muito mais abundantes em zoósporos móveis do que em células germinantes. Uma é uma tioredoxina, chamada TrxA, e a outra é sua proteína parceira, nomeada PtxA. Quando os pesquisadores deletaram os genes de TrxA ou de PtxA, os esporos ainda se formaram normalmente dentro dos esporângios e foram liberados no momento esperado—mas não nadaram. Microscopia eletrônica mostrou por quê: a maioria dos esporos desses mutantes simplesmente não possuía flagelos, ou tinha apenas alguns curtos. Ainda assim, os níveis de RNA mensageiro dos genes flagelares e a quantidade de pelo menos um bloco de construção proteico chave do flagelo (FliC) estavam essencialmente normais. Isso significa que TrxA e PtxA não ligam ou desligam genes flagelares; em vez disso, são necessárias para a montagem efetiva das estruturas do flagelo.
Uma função não redox para uma proteína clássica de redox
Tioredoxinas tipicamente atuam usando um par de aminoácidos cisteína para rearranjar ligações dissulfeto em outras proteínas. Para testar se essa atividade redox clássica era necessária aqui, a equipe purificou TrxA e mostrou, em um ensaio em tubo de ensaio, que ela se comporta como uma tioredoxina normal: pode reduzir um substrato proteico padrão. Quando mudaram uma ou ambas as cisteínas-chave, TrxA perdeu essa habilidade redox. Surpreendentemente, bactérias que carregavam essas versões “mortas para redox” de TrxA ainda produziram esporos totalmente flagelados e móveis. Em contraste, substituir parte de TrxA pela tioredoxina mais semelhante de outra bactéria não resgatou a formação de flagelos, mesmo que a proteína substituta tivesse atividade tioredox normal. Ao trocar sistematicamente regiões entre as duas proteínas, os autores refinaram a característica crucial até um curto trecho de cinco aminoácidos em TrxA, com a sequência EKVEQ, que é conservado entre muitas espécies de Actinoplanes.
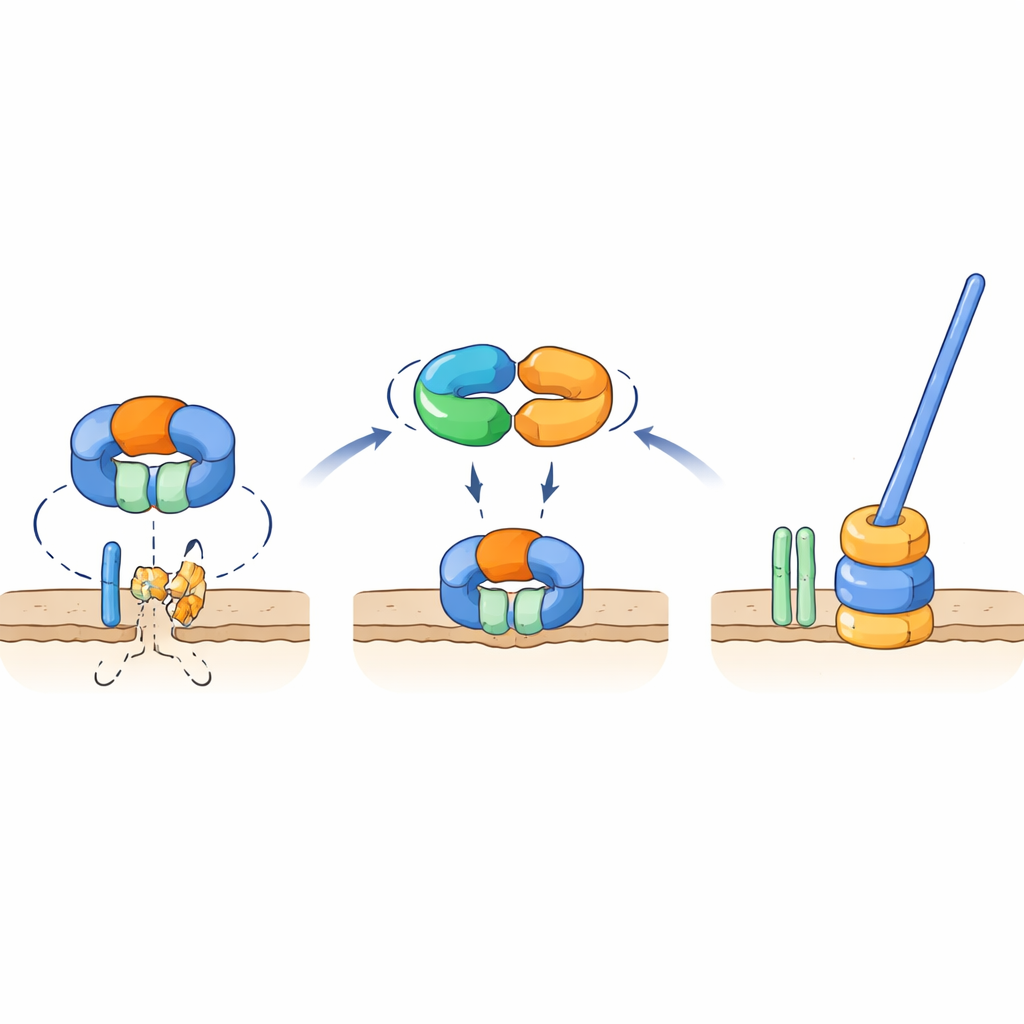
Um interruptor molecular que protege uma peça-chave do flagelo
Testes genéticos e de interação mostraram que TrxA e PtxA se ligam fisicamente entre si, e que o motivo EKVEQ é essencial para essa parceria. Usando um sistema bacteriano de two-hybrid, os pesquisadores descobriram que TrxA e PtxA também interagem com ClpC, um componente chaperona do complexo protease Clp—uma máquina molecular que desenrola e alimenta proteínas em um “triturador” em forma de barril. Para investigar como isso se relaciona com os flagelos, expuseram os mutantes não móveis de TrxA e PtxA à luz UV e selecionaram cepas supressoras raras cujos esporos recuperaram a motilidade. Muitas dessas supressoras apresentavam mutações em ClpC ou em FliR, uma proteína de membrana que faz parte do portão de exportação flagelar na base do apêndice. Introduzir essas mesmas mutações em cepas que careciam de TrxA ou PtxA restaurou a motilidade, e deletar fliR em um fundo genético normal aboliu completamente os flagelos. Esses achados sustentam um modelo no qual, em condições não favoráveis à formação de flagelos, a protease contendo ClpC degrada FliR, impedindo a montagem da base flagelar. Quando as condições favorecem a formação de flagelos, o complexo TrxA–PtxA liga-se a ClpC, atenuando sua atividade proteolítica para que FliR possa se acumular e o portão de exportação—e então o flagelo completo—possa ser construído.
Por que isso importa para a vida microbiana e a evolução de proteínas
Este trabalho revela um sistema finamente ajustado que permite a uma bactéria alternar rapidamente entre um esporo dormente e um nadador ativo protegendo um único componente vulnerável da máquina flagelar. Também mostra uma tioredoxina desempenhando uma função que não depende de sua química habitual: em vez de atuar como catalisador redox, TrxA usa um pequeno motivo conservado para formar um complexo regulatório com PtxA e controlar uma protease. Esse tipo de troca de função destaca como famílias de proteínas existentes podem ser reaproveitadas durante a evolução em novos módulos regulatórios, permitindo que bactérias como A. missouriensis coordenem transições complexas do ciclo de vida com apenas um punhado de interações moleculares cuidadosamente posicionadas.
Citação: Kimura, T., Maeda, S., Suzuki, R. et al. Thioredoxin and its partner protein are essential for zoospore flagellar formation in Actinoplanes missouriensis. Commun Biol 9, 532 (2026). https://doi.org/10.1038/s42003-026-09784-8
Palavras-chave: motilidade bacteriana, montagem do flagelo, regulação de proteínas, tioredoxina, controle por protease