Clear Sky Science · es
Tioredoxina y su proteína compañera son esenciales para la formación del flagelo de zoosporas en Actinoplanes missouriensis
Cómo algunas bacterias construyen colas diminutas a demanda
Muchas bacterias se desplazan usando largas colas parecidas a látigos llamadas flagelos. Para un microbio del suelo llamado Actinoplanes missouriensis, construir estas colas en el momento justo es cuestión de supervivencia: sus esporas deben volverse nadadoras de repente cuando aparece lluvia o agua. Este artículo desvela cómo un par de proteínas pequeñas actúa como un interruptor molecular de encendido y apagado para controlar cuándo se construyen esas colas nadadoras.
De esporas silenciosas a nadadoras repentinas
A. missouriensis normalmente crece como filamentos ramificados en el suelo. En condiciones secas, forma estructuras redondeadas llamadas esporangios en las puntas de esos filamentos. Dentro de cada esporangio se forman cientos de esporas redondas que se preparan para la vida exterior. Cuando llega el agua, el esporangio se abre y libera las esporas al medio líquido. Estas esporas brotan rápidamente flagelos y se convierten en «zoosporas» que pueden nadar durante un corto tiempo, ayudándolas a dispersarse a nuevos lugares antes de asentarse y volver a crecer. Dado que esta fase de natación es breve y está finamente cronometrada, la célula debe controlar con precisión cuándo y cómo se ensamblan los flagelos.
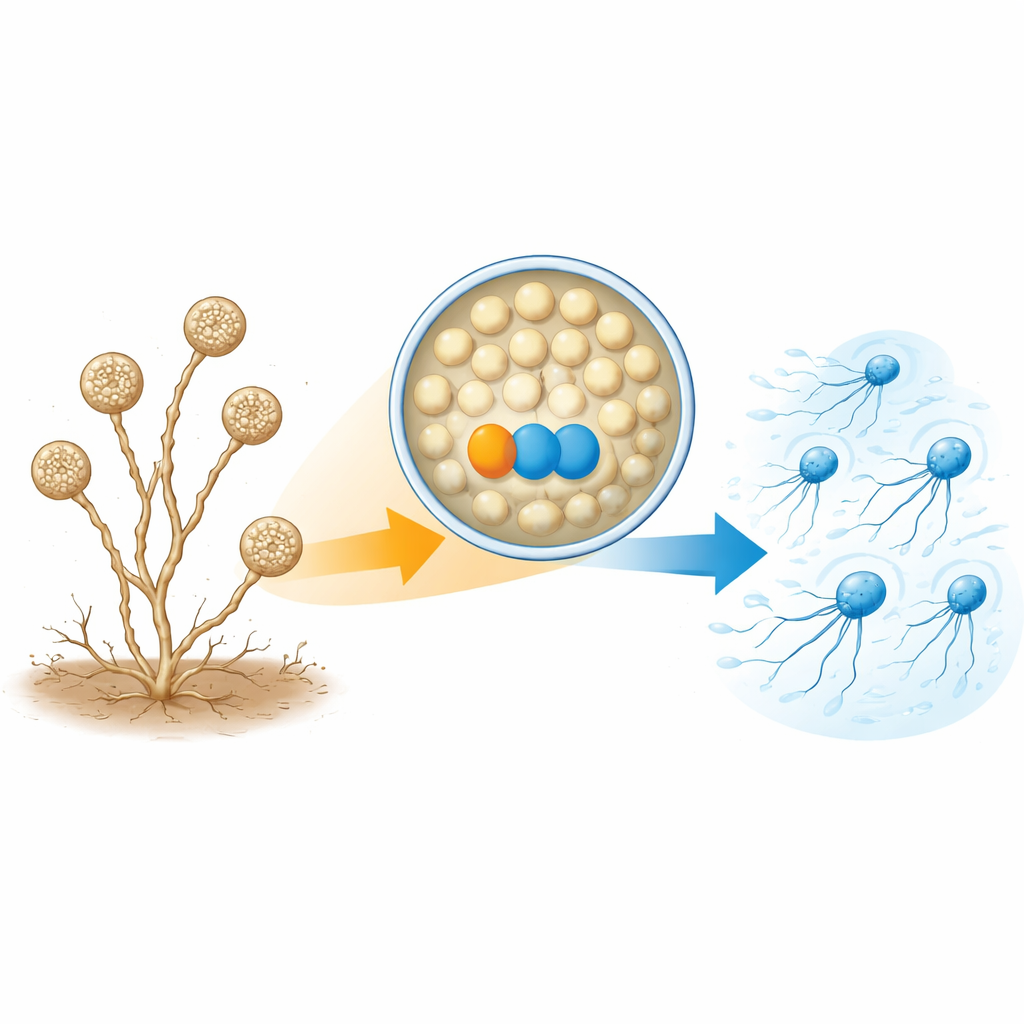
Una pareja de proteínas necesaria para construir colas
Los autores se centraron en dos proteínas que son mucho más abundantes en zoosporas nadadoras que en células en germinación. Una es una tioredoxina, denominada TrxA, y la otra es su proteína compañera, llamada PtxA. Cuando los investigadores eliminaron los genes de TrxA o PtxA, las esporas seguían formándose con normalidad dentro de los esporangios y se liberaban a tiempo, pero no nadaban. La microscopía electrónica mostró por qué: la mayoría de las esporas de estos mutantes simplemente carecían de flagelos, o tenían solo algunos cortos. Sin embargo, los niveles de ARN mensajero de los genes flagelares y la cantidad de al menos un bloque de construcción clave del flagelo (FliC) eran esencialmente normales. Esto indica que TrxA y PtxA no activan o desactivan los genes flagelares; en cambio, son necesarios para el ensamblaje real de las estructuras flagelares.
Un trabajo no redox para una proteína clásica de redox
Las tioredoxinas suelen actuar usando un par de residuos de cisteína para reorganizar enlaces disulfuro en otras proteínas. Para probar si esta actividad redox clásica era necesaria aquí, el equipo purificó TrxA y mostró en un ensayo in vitro que se comporta como una tioredoxina normal: puede reducir un sustrato proteico estándar. Cuando cambiaron una o ambas cisteínas clave, TrxA perdió esa capacidad redox. Sorprendentemente, las bacterias que portaban estas versiones «muertas para redox» de TrxA seguían produciendo esporas totalmente flageladas y móviles. En contraste, sustituir parte de TrxA por la tioredoxina más similar de otra bacteria no rescató la formación de flagelos, a pesar de que la proteína sustituta tenía actividad tioredox normal. Al intercambiar sistemáticamente regiones entre las dos proteínas, los autores acotaron la característica crucial a un tramo corto de cinco aminoácidos en TrxA, con la secuencia EKVEQ, que está conservado en muchas especies de Actinoplanes.
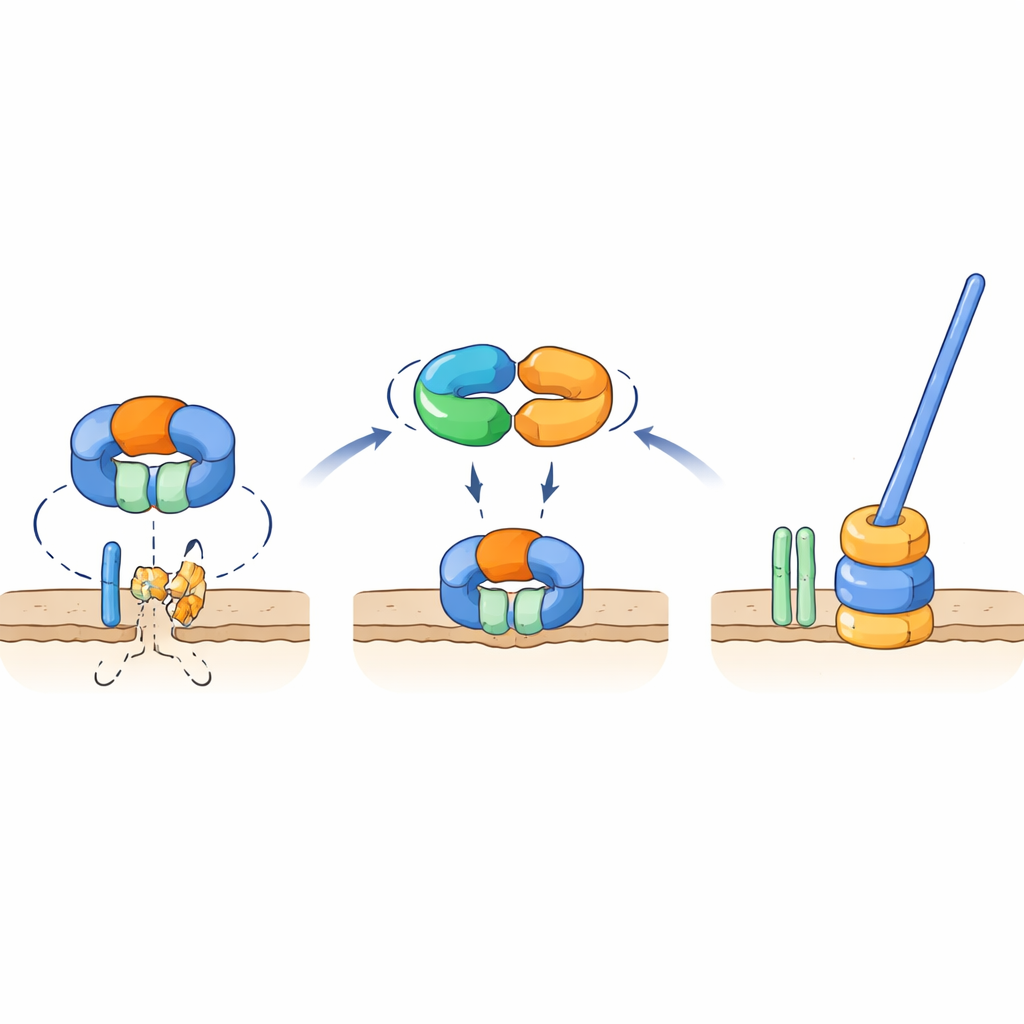
Un interruptor molecular que protege una pieza clave del flagelo
Pruebas genéticas y de interacción mostraron que TrxA y PtxA se unen físicamente entre sí, y que el motivo EKVEQ es esencial para esta asociación. Usando un sistema bacteriano de dos híbridos, los investigadores encontraron que TrxA y PtxA también interactúan con ClpC, un componente chaperón del complejo proteasa Clp—una máquina molecular que desenrolla y alimenta proteínas hacia un «triturador» con forma de barril. Para indagar cómo se relaciona esto con el flagelo, expusieron los mutantes no móviles de TrxA y PtxA a luz UV y seleccionaron cepas supresoras raras cuyas esporas recuperaron la motilidad. Muchas de estas supresoras portaban mutaciones en ClpC o en FliR, una proteína de membrana que forma parte de la puerta de exportación flagelar en la base de la cola. Introducir esas mismas mutaciones en cepas carentes de TrxA o PtxA restauró la motilidad, y eliminar fliR en un fondo genético normal abolió por completo los flagelos. Estos hallazgos respaldan un modelo en el que, en condiciones no flageladas, la proteasa que contiene ClpC degrada FliR, impidiendo el ensamblaje de la base flagelar. Cuando las condiciones favorecen la formación de flagelos, el complejo TrxA–PtxA se une a ClpC, atenuando su actividad proteolítica para que FliR pueda acumularse y la puerta de exportación—y luego el flagelo completo—puedan construirse.
Por qué importa esto para la vida microbiana y la evolución de las proteínas
Este trabajo revela un sistema finamente ajustado que permite a una bacteria cambiar rápidamente entre una espora dormida y una nadadora activa protegiendo un único componente vulnerable de la maquinaria flagelar. También muestra una tioredoxina desempeñando una función que no depende de su química habitual: en lugar de actuar como catalizador redox, TrxA usa un motivo corto y conservado para formar un complejo regulador con PtxA y controlar una proteasa. Ese tipo de cambio de función subraya cómo familias proteicas existentes pueden reutilizarse durante la evolución como nuevos módulos regulatorios, permitiendo a bacterias como A. missouriensis coordinar transiciones complejas del ciclo vital con apenas unas cuantas interacciones moleculares colocadas con precisión.
Cita: Kimura, T., Maeda, S., Suzuki, R. et al. Thioredoxin and its partner protein are essential for zoospore flagellar formation in Actinoplanes missouriensis. Commun Biol 9, 532 (2026). https://doi.org/10.1038/s42003-026-09784-8
Palabras clave: motilidad bacteriana, ensamblaje del flagelo, regulación proteica, tioredoxina, control por proteasa