Clear Sky Science · zh
高阶Gephyrin寡聚体的冷冻电镜结构揭示抑制性突触后支架组织的原理
大脑制动如何保持平衡
我们的脑功能依赖于“前进”和“停止”信号之间的微妙平衡。由抑制性突触传递的“停止”信号可防止与癫痫、焦虑及其他疾病相关的异常放电。本研究聚焦于gephyrin——一种在显微水平上构建并固定抑制性受体的关键蛋白。通过冷冻电子显微镜可视化gephyrin的三维构象,作者揭示了该蛋白如何自组装为更大的结构,并以出人意料的精确度组织受体。
抑制性突触的无声建筑师
在许多抑制性突触中,gephyrin 是主要的组织者,固定两类受体:甘氨酸受体和 GABAA 受体。每个 gephyrin 分子具有刚性的末端结构域,由一段柔性的中间区连接,而不同的可变剪接体又增加了复杂性。多年来,科学家普遍认为全长 gephyrin 主要形成三聚体,并以此描绘出类似晶格的突触后支架结构。然而,近期对完整脑组织的成像提示了一种更灵活的受体与支架网状结构,表明以三聚体为中心的旧观念并不完整。
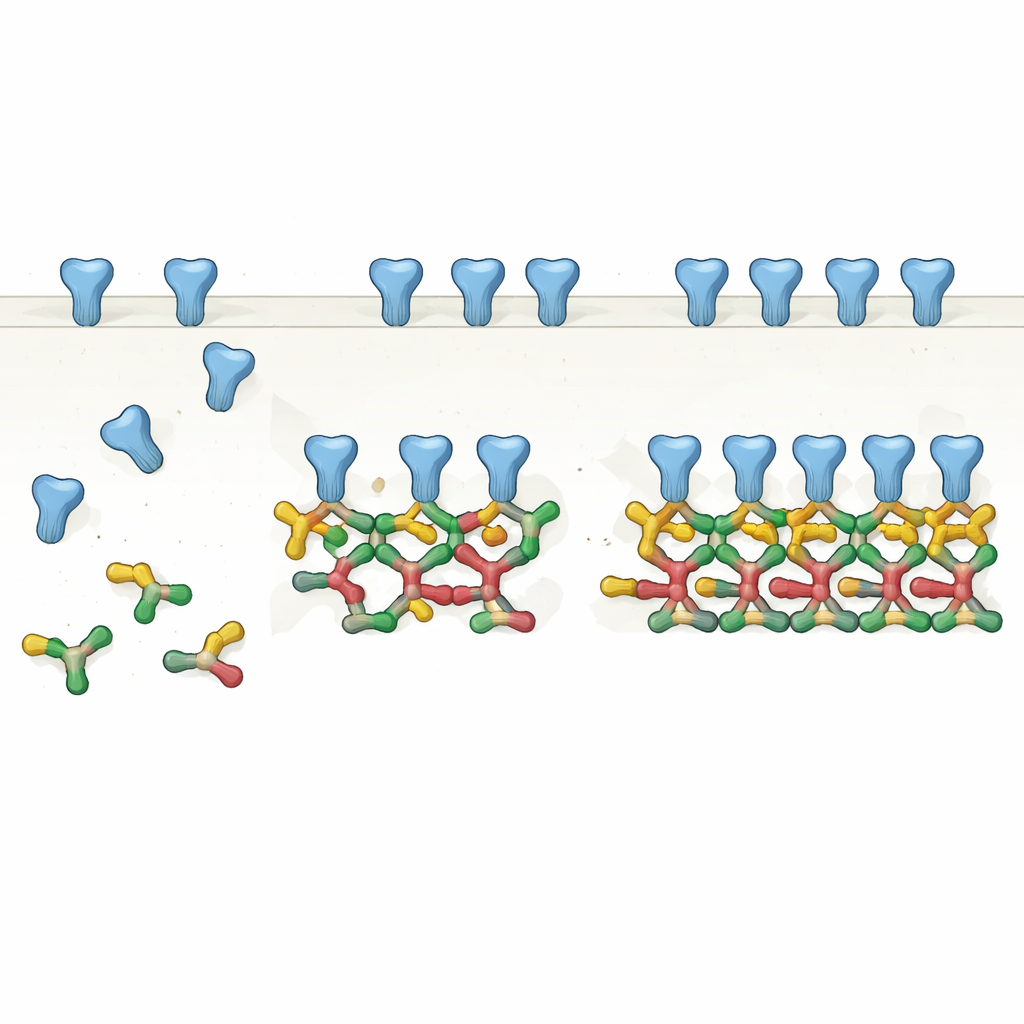
从对到链:对支架的新认识
通过精心纯化一种常见的全长 gephyrin 剪接体并用多种生化方法分析,作者表明 gephyrin 最自然的组装状态是成对的,即二聚体。这些二聚体随后作为基本构件进一步组装成更大结构:由四个分子组成的直线链(即二聚体的二聚体),甚至六聚体链。借助冷冻电子显微镜,他们解析了这些组装体的高分辨率结构,发现 gephyrin 某一部分的特定表面反复与相邻二聚体的匹配表面接触。与其说它是一个静态的三聚中心,不如说 gephyrin 是一个模块化系统,能够延伸成线性或成角的链条,从而匹配先前组织内成像中观察到的受体间距和排列。
带有隐蔽开关的柔性连接区
最令人着迷的发现之一在于蛋白的柔性中间段或连接区,该部分长期难以得到结构学解析。研究团队捕捉到该连接区回折并覆在 gephyrin 的一端结构域上,既稳定了蛋白,又延伸到用于结合受体的同一表面。在一种构象中,连接区的一部分位于受体结合口袋内,有效地阻塞了结合位点;在另一种构象中,它摆出位于外侧,让位点对受体开放。若干在“内/外”两种状态间改变位置的氨基酸是已知的磷酸化位点——磷酸化是细胞用作开关的常见化学修饰。这提示大脑细胞可能通过化学修饰将 gephyrin 的连接区在阻断受体和欢迎受体的构象之间切换,从而调节抑制强度。
静电“粘合剂”和液滴凝聚
研究还锁定了带电残基,它们充当静电“魔术贴”,帮助 gephyrin 二聚体连接成更高阶的链,并促进细胞内类液体蛋白液滴或凝聚体的形成。当研究者突变关键的正电或负电残基时,培养细胞中的 gephyrin 失去了形成大型液滴的能力,组装成的簇明显减少或更小。在神经元中,相同的突变削弱了 gephyrin 在抑制性突触处的富集,尽管被改变的蛋白仍能到达这些位点。这些实验共同表明,那些稳定 gephyrin 结构的带电区域和连接区,对在真实突触处形成致密的受体簇也是必不可少的。
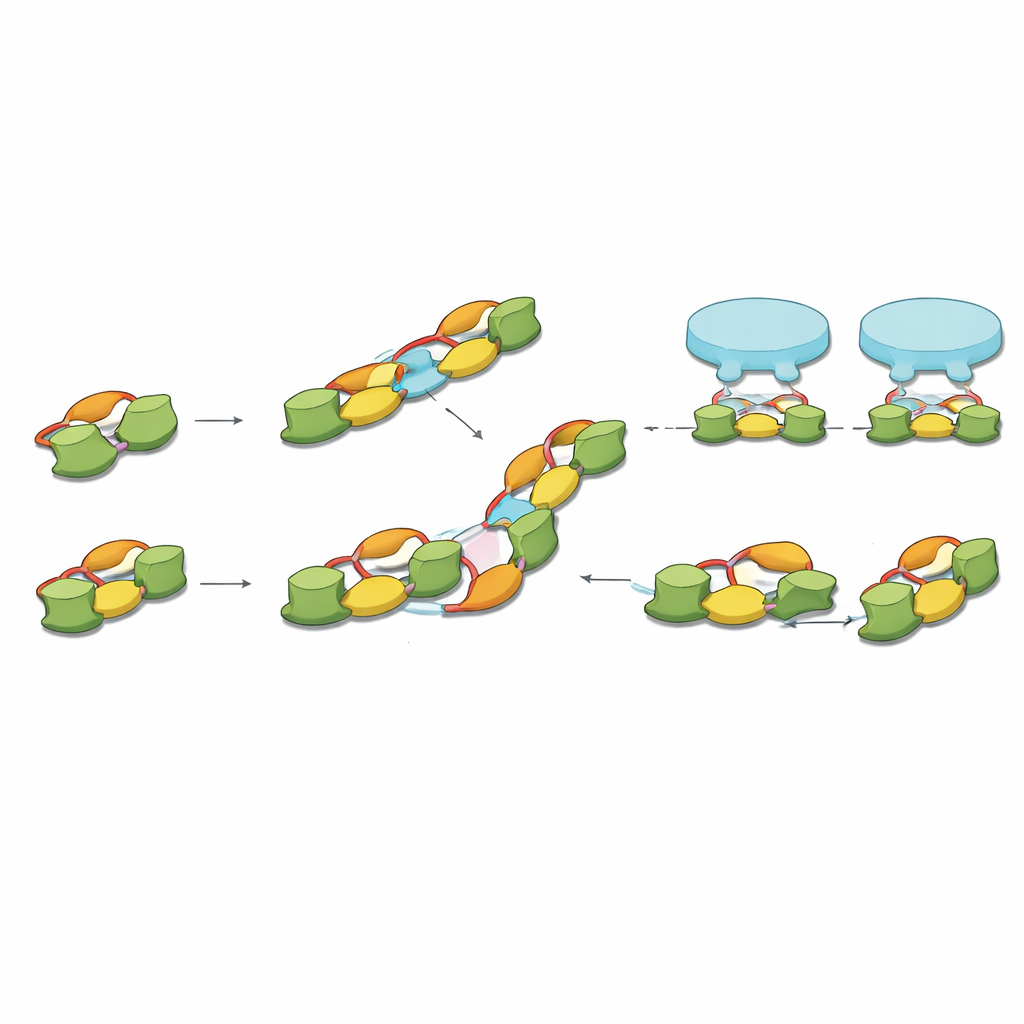
将分子联系到突触格局
最后,作者将他们的分子快照与早期对完整脑组织的冷冻电子断层成像联系起来,该研究测量出了相邻抑制性受体之间的特征性间距。gephyrin 二聚体的二聚体长度与该间距非常吻合,而直链与成角链的组合可以重建在神经元中观察到的最常见受体模式。在这一图景中,gephyrin 二聚体为受体对提供基本的夹持装置,而高阶链则通过带电界面并排连接这些夹持器而形成。
这对大脑健康为何重要
总体而言,这项工作用一种动态的链形成支架取代了简单的三聚体示意图,其形状和受体结合能力可通过柔性连接区上的细微化学修饰进行调节。这有助于解释抑制性突触如何既具有结构秩序又能快速可塑,并为理解破坏 gephyrin 组装的疾病相关突变提供了分子框架。通过阐明大脑制动系统的物理构建和调节方式,该研究为在癫痫、自闭谱系障碍和焦虑等疾病中调节抑制性平衡开辟了新的思路。
引用: Ortiz-López, D., Hove, T.T., Huhn, C. et al. Cryo-EM structures of higher order Gephyrin oligomers reveal principles of inhibitory postsynaptic scaffold organization. Nat Commun 17, 3541 (2026). https://doi.org/10.1038/s41467-026-71771-8
关键词: gephyrin, 抑制性突触, GABAA 受体, 冷冻电子显微镜, 突触支架