Clear Sky Science · zh
代谢氧化还原酶:干性表观基因组的核心调控因子
细胞燃料如何塑造细胞身份
体内每个细胞携带的基本上都是相同的DNA,然而神经元、血细胞和皮肤细胞在外观和功能上却截然不同。本文探讨了细胞如何处理燃料与其决定成为什么类型细胞之间的一个令人意外的联系。文章解释了一类称为氧化还原酶的酶——它们通常处理能量与营养物质——如何同时参与在DNA和蛋白质上写入与擦除化学标记,这些标记控制哪些基因被激活。理解这一联系可能会重塑我们对干细胞、衰老乃至癌症的认知。
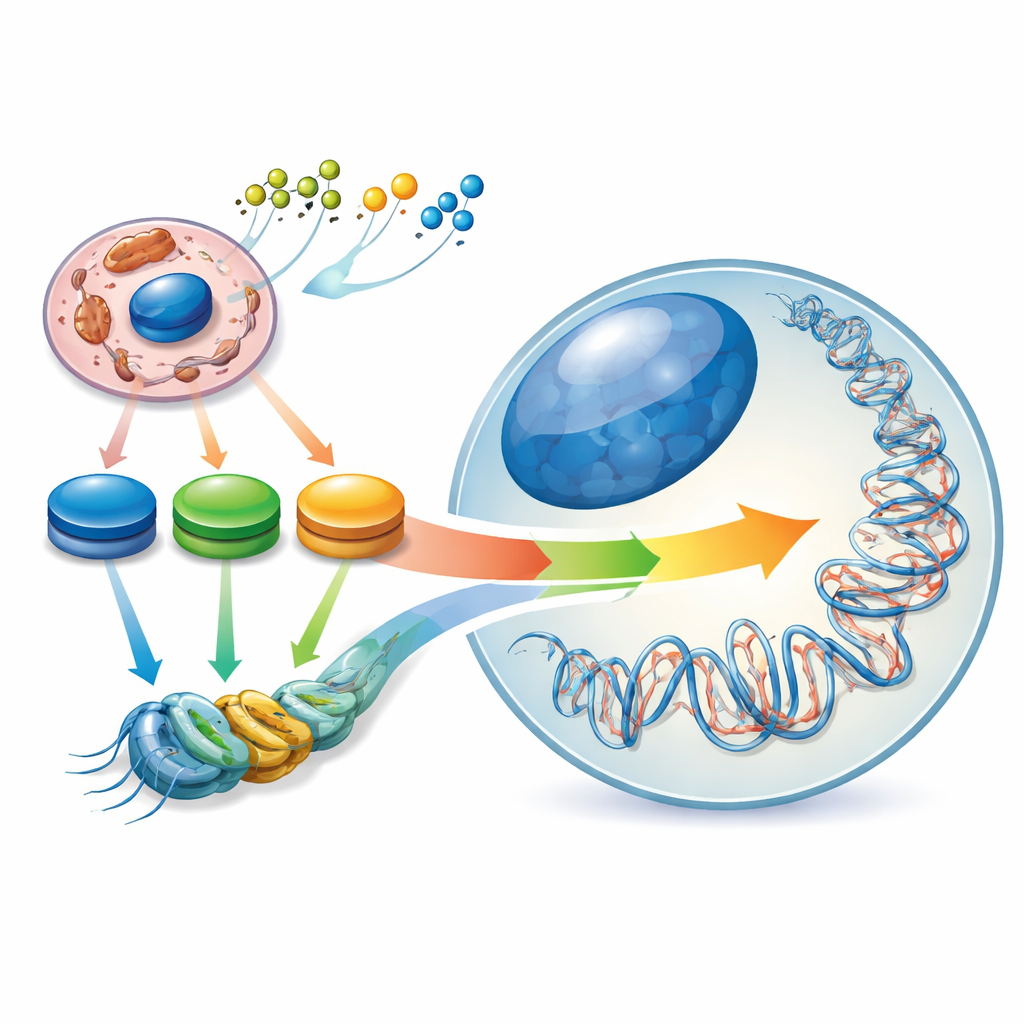
我们基因内部的隐秘景观
干细胞之所以特殊,是因为它们既能自我更新,又能分化为多种不同细胞类型。它们通过一种内部的“表观遗传景观”来维持这一平衡——即DNA及其包装蛋白上的化学标签模式,像调光开关一样调节基因。作者将这一景观描述为由细胞内的“能量等高线”塑造:关键代谢分子(如乙酰辅酶A、NAD⁺/NADH 和α-酮戊二酸)浓度的高低形成峰谷。当干细胞从幼稚状态转向更为承诺的状态时,其中心碳代谢会被重编程,表观遗传景观也随之改变。因此,细胞燃料利用的变化不仅是细胞身份的结果——它们也积极参与决定细胞身份。
将食物转化为表观遗传信号的酶
氧化还原酶催化氧化还原反应,处于代谢的关键枢纽。许多此类酶生成“表观代谢物”,这些分子作为标记DNA和组蛋白的酶的底物或辅因子。例如,糖酵解和三羧酸循环中的脱氢酶产生用于组蛋白乙酰化的乙酰辅酶A,产生影响去乙酰酶活性的NAD⁺/NADH和NADPH,以及驱动DNA和组蛋白去甲基化的α-酮戊二酸。通过控制这些代谢物的可用量,氧化还原酶设定了可能的表观遗传变化范围。不同的干细胞状态依赖不同的代谢组合:幼稚的多能干细胞同时利用糖类和脂肪,准备状态的多能细胞强烈依赖糖酵解和谷氨酰胺,许多成人干细胞则偏向脂肪酸分解和氧化磷酸化。
直接编辑DNA、RNA与组蛋白的酶
部分氧化还原酶不仅提供原料,还直接重写表观遗传标记。依赖铁和α-酮戊二酸的酶,如TET蛋白和JMJD去甲基酶,可以从DNA和组蛋白上去除甲基,重塑基因的活性。相关酶ALKBH5和FTO能擦除RNA上的甲基标记,影响mRNA的稳定性和翻译效率。另一类酶LSD1和LSD2利用黄素辅因子去甲基化特定的组蛋白赖氨酸残基。由于这些酶都依赖代谢辅因子和氧气,它们的活性对细胞的能量和氧化还原状态极为敏感。综述展示了α-酮戊二酸、琥珀酸或癌症中突变代谢物的变化,如何将开放且可塑的染色质与紧闭受抑制的状态之间的平衡撬动过去。
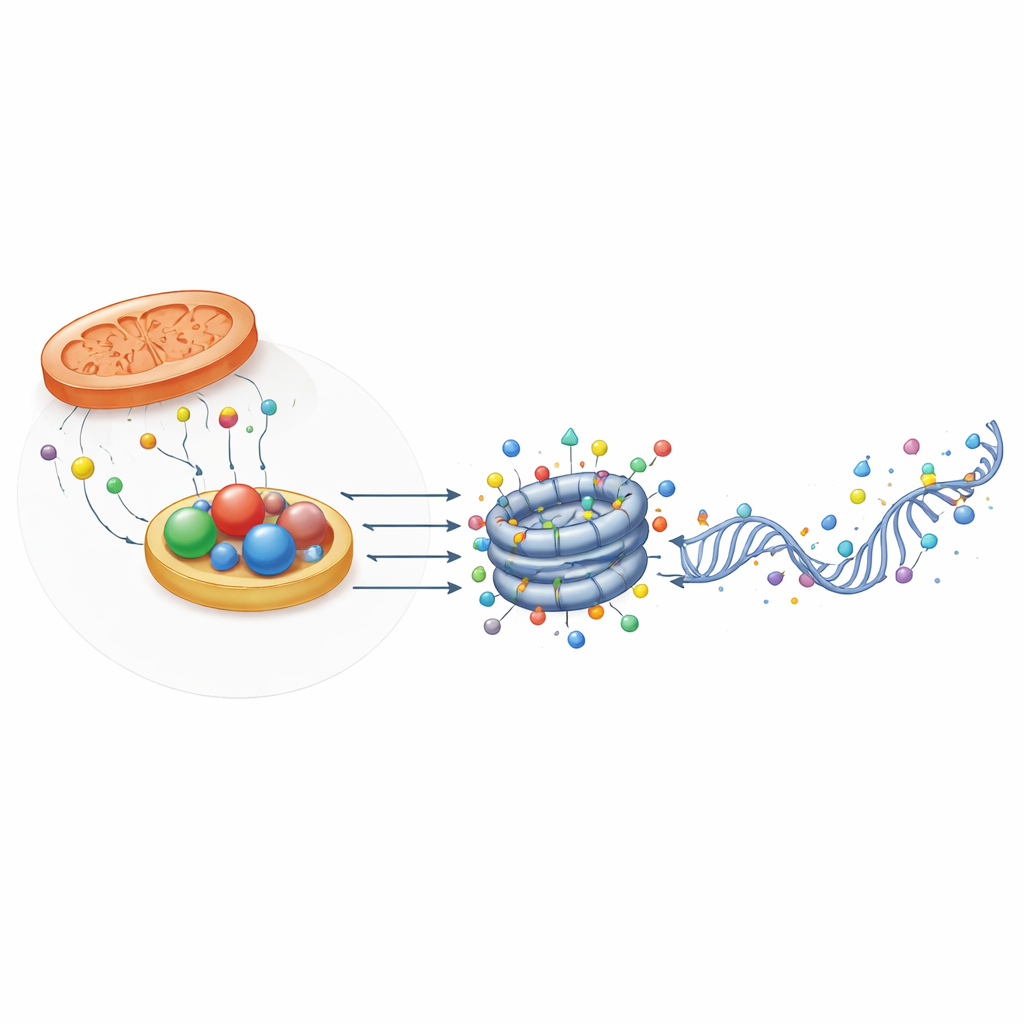
感知氧化还原状态的结构性助力蛋白
耐人寻味的是,并非所有与氧化还原相关的蛋白都作为酶发挥作用。有些蛋白,如CTBP和NPAC/GLYR1,保留了能结合NADH的结构折叠但几乎没有催化活性。相反,它们形成多单元支架,将转录因子和染色质修饰因子带到特定基因区域。它们的组装依赖于NAD(H)结合,这意味着它们可以作为细胞氧化还原状态的物理传感器。在小鼠胚胎干细胞中,CTBP2通过招募去乙酰和去甲基复合体,帮助使多能基因在分化时易于关闭;而NPAC/GLYR1则与LSD2和RNA聚合酶II协同,协调转录延伸和可变剪接。这些非酶学功能为代谢如何影响基因调控再添一层机制。
这对健康与疾病的重要性
作者总结道,氧化还原酶在三个层面上将代谢与基因调控串联起来:它们生成关键代谢物,直接擦除DNA、RNA和组蛋白上的化学标记,以及作为支架重组染色质机制。合在一起,这些功能把细胞的燃料使用转化为关于应表达哪些基因的持久指令,帮助维持干性或推动分化。这一框架有助于解释为何代谢相关突变能将细胞重编程为类似癌症的状态,以及为何调整营养或代谢酶能改变干细胞行为。随着研究者更细致地探究这些联系,氧化还原酶正成为有吸引力的再生疗法和抗癌策略靶点,旨在通过重塑细胞处理能量的方式来改造表观遗传景观。
引用: Lee, HT., Roe, JS. & Youn, HD. Metabolic oxidoreductases: central regulators of the epigenetic landscapes in stemness. Exp Mol Med 58, 1017–1037 (2026). https://doi.org/10.1038/s12276-026-01687-2
关键词: 干细胞代谢, 表观遗传调控, 氧化还原酶, 染色质重塑, 癌症干细胞