Clear Sky Science · sv
Metabola oxidoreduktaser: centrala regulatorer av det epigenetiska landskapet i stamcellers egenskaper
Hur cellens bränsle formar cellidentitet
Varje cell i din kropp bär i grunden samma DNA, ändå ser neuroner, blodceller och hudceller helt olika ut och beter sig olika. Denna artikel undersöker en oväntad koppling mellan hur celler bearbetar bränsle och hur de bestämmer vilken celltyp de ska bli. Den förklarar hur en klass enzymer kallade oxidoreduktaser, som normalt hanterar energi och näringsämnen, också hjälper till att skriva och radera de kemiska markörer på DNA och proteiner som styr vilka gener som är aktiva. Att förstå denna koppling kan förändra hur vi ser på stamceller, åldrande och till och med cancer.
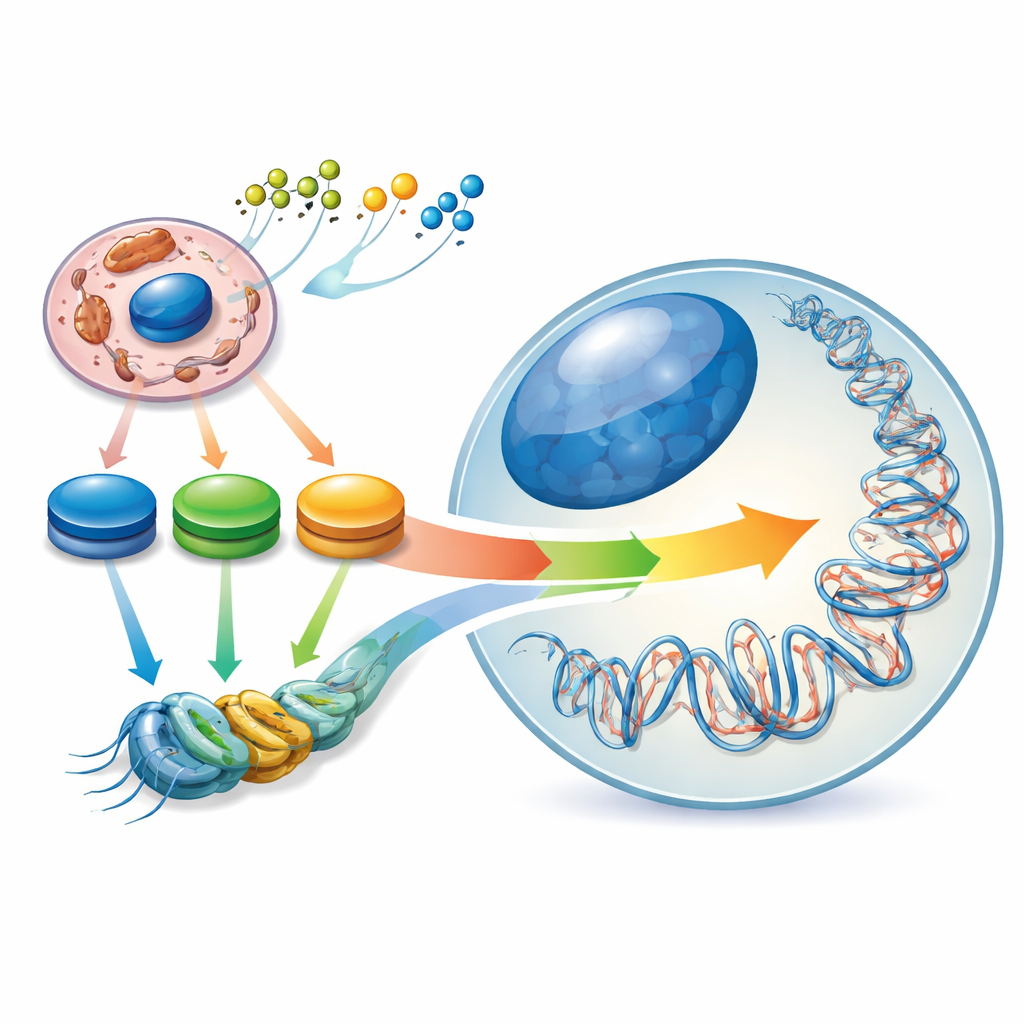
Det dolda landskapet inne i våra gener
Stamceller är speciella eftersom de antingen kan förnya sig själva eller mogna till många olika celltyper. De balanserar detta genom ett internt ”epigenetiskt landskap” – mönster av kemiska taggar på DNA och dess packningsproteiner som fungerar som dimmers för gener. Författarna beskriver detta landskap som format av en ”energi-konturnivå” inne i cellen: toppar och dalar i nivåerna av nyckelmetaboliska molekyler som acetyl-CoA, NAD⁺/NADH och α-ketoglutarat. När en stamcell skiftar från ett naivt tillstånd till ett mer åtagit skiftas också dess centrala kolmetabolism, och det epigenetiska landskapet följer med. Förändringar i hur cellen använder bränsle är alltså inte bara konsekvenser av cellidentitet – de hjälper aktivt till att bestämma den.
Enzymer som förvandlar mat till epigenetiska signaler
Oxidoreduktaser är enzymer som katalyserar oxidations–reduktionsreaktioner och sitter i nyckelknutpunkter i metabolismen. Många av dem producerar ”epigenetiska metaboliter”, molekyler som tjänar som råmaterial eller kofaktorer för enzymer som sätter taggar på DNA och histoner. Exempelvis producerar dehydrogenaser i glykolysen och TCA-cykeln acetyl-CoA för histonacetylering, NAD⁺/NADH och NADPH som påverkar deacetylaser, och α-ketoglutarat som driver DNA- och histondemetylaser. Genom att kontrollera hur mycket av dessa metaboliter som finns tillgängliga bestämmer oxidoreduktaserna spannet av möjliga epigenetiska förändringar. Olika stamcellstillstånd förlitar sig på olika metaboliska blandningar: naiva pluripotenta stamceller utnyttjar både socker och fett, primade pluripotenta celler lutar starkt mot glykolys och glutamin, och många vuxna stamceller föredrar fettsyreoxidation och oxidativ fosforylering.
Direkta redigerare av DNA, RNA och histoner
Vissa oxidoreduktaser gör mer än att bara leverera ingredienser — de skriver direkt om epigenetiska markörer. Järn- och α-ketoglutarat‑beroende enzymer såsom TET-proteiner och JMJD-demetylaser tar bort metylgrupper från DNA och histoner och omformar vilka gener som är aktiva. Besläktade enzymer, ALKBH5 och FTO, raderar metylmarkeringar på RNA, vilket påverkar hur länge budskap varar och hur effektivt de översätts. En annan grupp, LSD1 och LSD2, använder en flavin-kofaktor för att demetylsera specifika histonlysinrester. Eftersom alla dessa enzymer kräver metabola kofaktorer och syre är deras aktivitet mycket känslig för cellens energi- och redoxstatus. Översikten visar hur skiften i α-ketoglutarat, succinat eller mutanta metaboliter i cancer kan rubba balansen mellan öppet, flexibelt kromatin och tätt låsta, repressiva tillstånd.
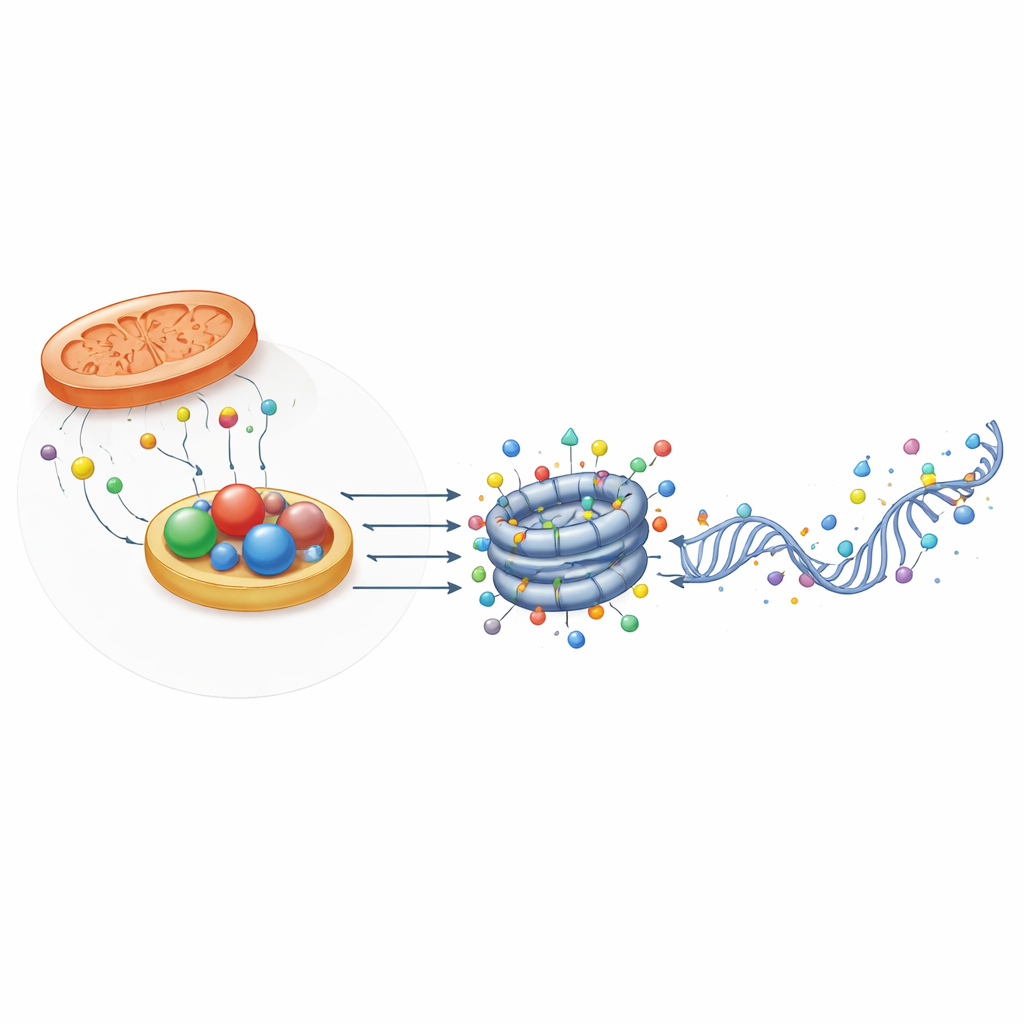
Strukturella hjälpare som känner av redoxstatus
Intressant nog fungerar inte alla oxidoreduktasrelaterade proteiner som enzymer. Vissa, såsom CTBP och NPAC/GLYR1, har kvar den strukturella veckningen som binder NADH men har liten eller ingen katalytisk aktivitet. Istället bildar de flerenhetsplattformar som samlar transkriptionsfaktorer och kromatinmodifierare vid specifika genregioner. Deras montering beror på NAD(H)-bindning, vilket innebär att de kan fungera som fysiska sensorer för cellens redoxstatus. I musens embryonala stamceller bidrar CTBP2 till att göra pluripotensgener redo att slås av under differentiering genom att rekrytera komplex som tar bort acetyl- och metylmärken, medan NPAC/GLYR1 samarbetar med LSD2 och RNA-polymeras II för att samordna transkriptionell elongation och alternativ splitsning. Dessa icke-enzymatiska roller lägger till ytterligare ett lager där metabolism påverkar genreglering.
Varför detta spelar roll för hälsa och sjukdom
Författarna drar slutsatsen att oxidoreduktaser knyter ihop metabolism och genreglering på tre nivåer: de genererar nyckelmetaboliter, raderar direkt kemiska markörer på DNA, RNA och histoner, och fungerar som scaffoldar som omorganiserar kromatinmaskineriet. Tillsammans förvandlar dessa funktioner cellens bränsleanvändning till bestående instruktioner om vilka gener som ska uttryckas, vilket hjälper till att bevara stamcellsegenskaper eller driva differentiering. Denna ramverk hjälper till att förklara varför metabola mutationer kan omprogrammera celler mot cancerliknande tillstånd, och varför justeringar av näringsämnen eller metabola enzymer kan förändra stamcellsbeteende. När forskare undersöker dessa länkar mer i detalj framstår oxidoreduktaser som attraktiva mål för regenerativa terapier och anticancerstrategier som syftar till att omforma det epigenetiska landskapet genom att omkoppla hur celler hanterar energi.
Citering: Lee, HT., Roe, JS. & Youn, HD. Metabolic oxidoreductases: central regulators of the epigenetic landscapes in stemness. Exp Mol Med 58, 1017–1037 (2026). https://doi.org/10.1038/s12276-026-01687-2
Nyckelord: stamcellsmetabolism, epigenetisk reglering, oxido-reduktasenzym, kromatinomformning, cancerstamceller