Clear Sky Science · pl
Oksydoreduktazy metaboliczne: centralni regulatorzy epigenetycznych krajobrazów w zachowaniu pluripotencji
Jak paliwo komórkowe kształtuje tożsamość komórki
Każda komórka w twoim ciele posiada w zasadzie to samo DNA, a mimo to neurony, krwinki i komórki skóry wyglądają i działają zupełnie inaczej. Artykuł bada zaskakujące powiązanie między tym, jak komórki przetwarzają paliwo, a tym, jak decydują, jakim typem komórki się staną. Wyjaśnia, jak klasa enzymów zwana oksydoreduktazami, które zwykle zajmują się energią i składnikami odżywczymi, pomaga też zapisywać i usuwać chemiczne znaczniki na DNA i białkach, które kontrolują, które geny są włączone. Zrozumienie tej zależności może zmienić nasze spojrzenie na komórki macierzyste, starzenie się, a nawet nowotwory.
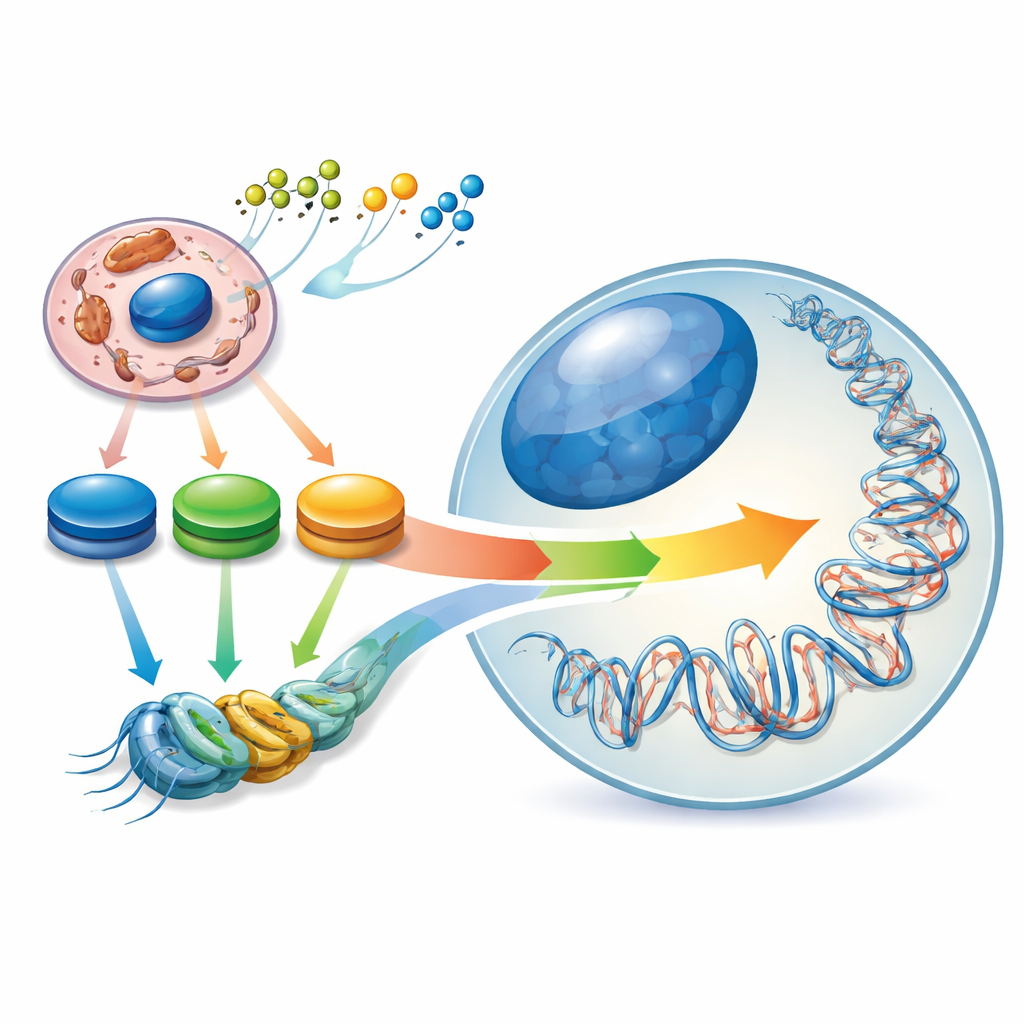
Ukryty krajobraz wewnątrz naszych genów
Komórki macierzyste są wyjątkowe, ponieważ mogą albo odnawiać samy siebie, albo dojrzewać w wiele różnych typów komórek. Utrzymują tę równowagę dzięki wewnętrznemu „epigenetycznemu krajobrazowi” – wzorom chemicznych znaczników na DNA i białkach pakujących chromatynę, które działają jak ściemniacze genów. Autorzy opisują ten krajobraz jako formowany przez wewnętrzny „kontur energetyczny”: szczyty i doliny w poziomach kluczowych cząsteczek metabolicznych, takich jak acetylo-CoA, NAD⁺/NADH i α‑ketoglutaran. Gdy komórka macierzysta przechodzi ze stanu naiwnych w kierunku bardziej zaangażowanego, jej centralny metabolizm węglowy jest przestawiany, a wraz z nim zmienia się krajobraz epigenetyczny. Zmiany w wykorzystaniu paliwa komórkowego nie są więc tylko następstwem tożsamości komórkowej – aktywnie pomagają ją określić.
Enzymy, które zamieniają pokarm w sygnały epigenetyczne
Oksydoreduktazy to enzymy katalizujące reakcje utleniania–redukcji i zajmujące kluczowe węzły metabolizmu. Wiele z nich wytwarza „metabolity epigenetyczne”, cząsteczki będące surowcem lub kofaktorami dla enzymów znakujących DNA i histony. Na przykład dehydrogenazy w glikolizie i cyklu TCA produkują acetylo‑CoA do acetylacji histonów, NAD⁺/NADH i NADPH wpływające na deacetylazy oraz α‑ketoglutaran napędzający demetylazy DNA i histonów. Kontrolując dostępność tych metabolitów, oksydoreduktazy wyznaczają zakres możliwych zmian epigenetycznych. Różne stany komórek macierzystych polegają na odmiennych mieszankach metabolicznych: naiwni pluripotentni korzystają i z cukrów, i z tłuszczów, spośród „primed” pluripotentnych dominują glikoliza i glutamina, a wiele dorosłych komórek macierzystych preferuje rozkład kwasów tłuszczowych i fosforylację oksydacyjną.
Bezpośredni redaktorzy DNA, RNA i histonów
Niektóre oksydoreduktazy robią więcej niż dostarczają składników – bezpośrednio przepisują znaki epigenetyczne. Enzymy zależne od żelaza i α‑ketoglutaranu, takie jak białka TET i demetylazy JMJD, usuwają grupy metylowe z DNA i histonów, przekształcając, które geny są aktywne. Powiązane enzymy, ALKBH5 i FTO, zmazują metylację na RNA, wpływając na długość życia cząsteczek informacyjnych i efektywność ich translacji. Inna grupa, LSD1 i LSD2, wykorzystuje kofaktor flawinowy do demetylacji wybranych lizyn histonowych. Ponieważ wszystkie te enzymy wymagają metabolicznych kofaktorów i tlenu, ich aktywność jest wysoce wrażliwa na stan energetyczny i redoks komórki. Przegląd pokazuje, jak zmiany stężeń α‑ketoglutaranu, bursztynianu lub mutantnych metabolitów w nowotworach mogą przesunąć równowagę między otwartą, elastyczną chromatyną a mocno zamkniętymi, zredukowanymi stanami.
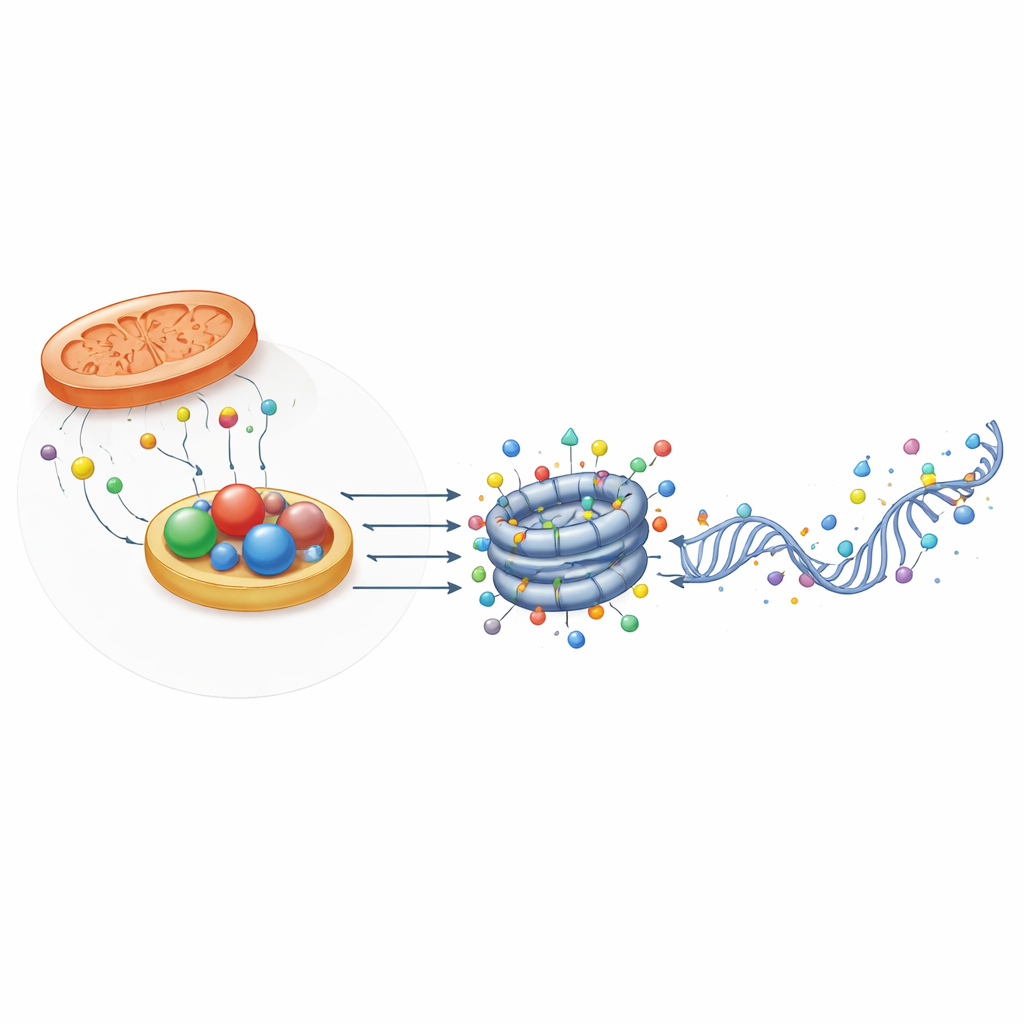
Strukturalne pomocniki wyczuwające stan redoks
Zaskakująco, nie wszystkie białka powiązane z oksydoreduktazami pełnią funkcje enzymatyczne. Niektóre, jak CTBP i NPAC/GLYR1, zachowują fałd strukturalny wiążący NADH, ale mają niewielką lub brak aktywności katalitycznej. Zamiast tego tworzą wielopodjednostkowe rusztowania, które łączą czynniki transkrypcyjne i modyfikatory chromatyny w określonych regionach genów. Ich składanie zależy od wiązania NAD(H), co oznacza, że mogą działać jako fizyczne czujniki stanu redoks komórki. W embrionalnych komórkach macierzystych myszy CTBP2 pomaga ustawić geny pluripotencji do wyłączenia podczas różnicowania przez rekrutację kompleksów usuwających acetyl i metyl, podczas gdy NPAC/GLYR1 współdziała z LSD2 i polimerazą RNA II, koordynując elongację transkryptu i alternatywne składanie. Te nieenzymatyczne role dodają kolejną warstwę, przez którą metabolizm wpływa na kontrolę genów.
Dlaczego to ma znaczenie dla zdrowia i chorób
Autorzy wnioskują, że oksydoreduktazy łączą metabolizm i regulację genów na trzech poziomach: wytwarzają kluczowe metabolity, bezpośrednio usuwają chemiczne znaczniki z DNA, RNA i histonów oraz pełnią rolę rusztowań reorganizujących aparat chromatynowy. Te funkcje sprawiają, że wykorzystanie paliwa przez komórkę przekształca się w trwałe instrukcje określające, które geny mają być wyrażone, pomagając utrzymać pluripotencję lub wymusić różnicowanie. Ta ramowa koncepcja pomaga wyjaśnić, dlaczego mutacje metaboliczne mogą przeprogramować komórki w kierunku stanów przypominających nowotwór oraz dlaczego modyfikacja składników odżywczych lub enzymów metabolicznych może zmieniać zachowanie komórek macierzystych. W miarę jak badacze zgłębiają te powiązania, oksydoreduktazy wyrastają na atrakcyjne cele dla terapii regeneracyjnych i przeciwnowotworowych, które dążą do przebudowy krajobrazu epigenetycznego poprzez przestawienie sposobu, w jaki komórki zarządzają energią.
Cytowanie: Lee, HT., Roe, JS. & Youn, HD. Metabolic oxidoreductases: central regulators of the epigenetic landscapes in stemness. Exp Mol Med 58, 1017–1037 (2026). https://doi.org/10.1038/s12276-026-01687-2
Słowa kluczowe: metabolizm komórek macierzystych, regulacja epigenetyczna, enzymy oksydoreduktazowe, remodeling chromatyny, komórki macierzyste nowotworów