Clear Sky Science · ru
Метаболические оксидоредуктазы: центральные регуляторы эпигенетического ландшафта стемнесса
Как топливо клетки формирует её идентичность
Каждая клетка вашего организма по сути несёт один и тот же набор ДНК, однако нейроны, кровяные клетки и клетки кожи выглядят и функционируют совершенно по-разному. В этой статье рассматривается неожиданная связь между тем, как клетка перерабатывает топливо, и тем, как она «решает», в какой тип клетки превратиться. Объясняется, как класс ферментов — оксидоредуктаз, которые обычно участвуют в обмене энергии и питательных веществ, — также помогают «писать» и «стирать» химические метки на ДНК и белках, контролирующие включение генов. Понимание этой связи может изменить наше представление о стволовых клетках, старении и даже раке.
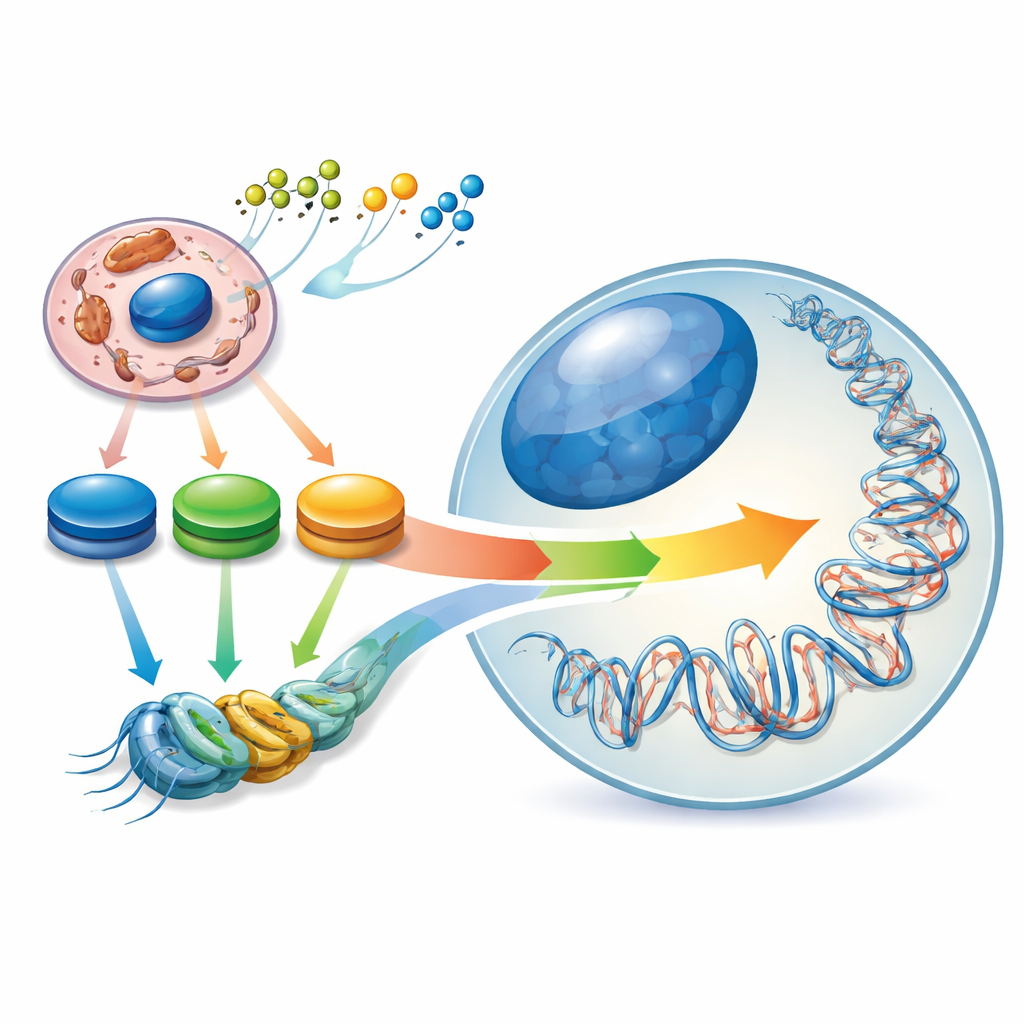
Скрытый ландшафт внутри наших генов
Стволовые клетки особенны тем, что они могут либо самоподдерживаться, либо дифференцироваться в многочисленные типы клеток. Они управляют этим балансом через внутренний «эпигенетический ландшафт» — узоры химических меток на ДНК и белках-упаковщиках, которые действуют как регуляторы интенсивности генов. Авторы описывают этот ландшафт как формируемый «энергетическим контуром» внутри клетки: пиков и впадин в уровнях ключевых метаболитов, таких как ацетил-CoA, NAD⁺/NADH и α-кетоглутарат. Когда стволовая клетка переходит из наивного состояния в более обусловленное, её центральный углеродный метаболизм перенастраивается, и эпигенетический ландшафт смещается вместе с ним. Таким образом, изменения в потреблении топлива клеткой — не просто следствие её идентичности, они активно помогают её определять.
Ферменты, превращающие пищу в эпигенетические сигналы
Оксидоредуктазы — это ферменты, катализирующие реакции окисления-восстановления и находящиеся в ключевых узлах метаболизма. Многие из них производят «эпигенетические метаболиты», молекулы, служащие сырьём или кофакторами для ферментов, наносящих метки на ДНК и гистоны. Например, дегидрогеназы в гликолизе и цикле Кребса вырабатывают ацетил-CoA для ацетилирования гистонов, NAD⁺/NADH и NADPH, влияющие на деацетилазы, а также α-кетоглутарат, питающий деметилазы ДНК и гистонов. Контролируя доступность этих метаболитов, оксидоредуктазы задают диапазон возможных эпигенетических изменений. Разные состояния стволовых клеток опираются на различные метаболические смеси: наивные плюрипотентные клетки используют и углеводы, и жиры, «приготовленные» (primed) плюрипотентные клетки сильно зависят от гликолиза и глутамина, а многие взрослые стволовые клетки предпочитают распад жирных кислот и окислительное фосфорилирование.
Прямые редакторы ДНК, РНК и гистонов
Некоторые оксидоредуктазы идут дальше простого снабжения ингредиентами и непосредственно переписывают эпигенетические метки. Железо- и α-кетоглутарат-зависимые ферменты, такие как белки TET и деметилазы семейства JMJD, удаляют метильные группы с ДНК и гистонов, изменяя активность генов. Родственные ферменты, ALKBH5 и FTO, стирают метки метилирования на РНК, влияя на стабильность молекул и эффективность их трансляции. Другая группа, LSD1 и LSD2, использует флавиновый кофактор для деметилирования определённых лизинов гистонов. Поскольку все эти ферменты требуют метаболических кофакторов и кислорода, их активность крайне чувствительна к энергетическому и редокс-состоянию клетки. Обзор показывает, как сдвиги в уровнях α-кетоглутарата, сукцината или появление мутантных метаболитов при раке могут сдвинуть баланс между открытым, гибким хроматином и туго «заблокированными», репрессированными состояниями.
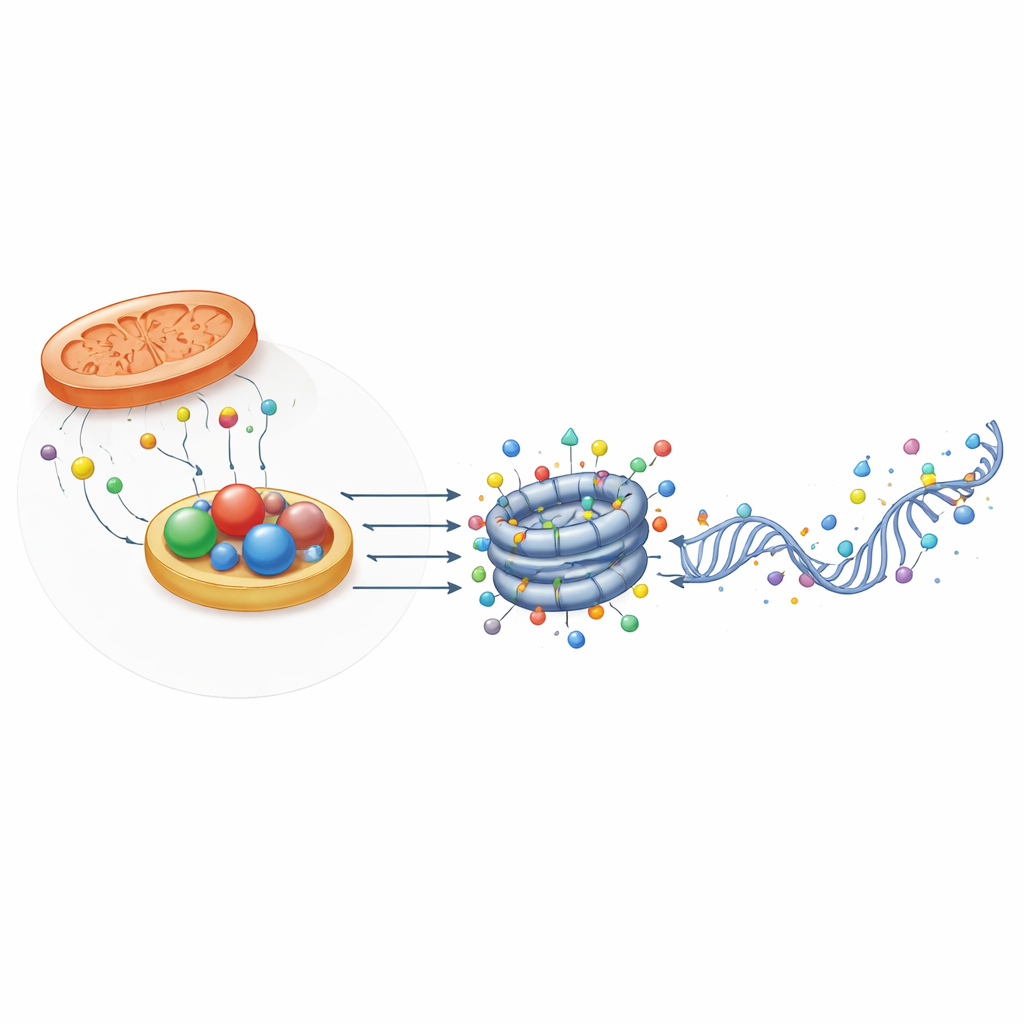
Структурные помощники, улавливающие редокс-состояние
Любопытно, что не все белки, связанные с оксидоредуктазами, действуют как ферменты. Некоторые, такие как CTBP и NPAC/GLYR1, сохранили структурный фолд, связывающий NADH, но практически лишены каталитической активности. Вместо этого они формируют многоузловые каркасы, которые сводят вместе факторы транскрипции и модификаторы хроматина в определённых регионах генома. Их сборка зависит от связывания NAD(H), что позволяет им выступать в роли физических сенсоров редокс-состояния клетки. В эмбриональных стволовых клетках мыши CTBP2 помогает подготавливать гены плюрипотентности к выключению при дифференцировке, привлекая комплексы, удаляющие ацетильные и метильные метки, в то время как NPAC/GLYR1 объединяется с LSD2 и РНК-полимеразой II для координацииelongation транскрипции и альтернативного сплайсинга. Эти неэнзимные роли добавляют ещё один уровень того, как метаболизм влияет на управление генами.
Почему это важно для здоровья и болезни
Авторы приходят к выводу, что оксидоредуктазы связывают метаболизм и регуляцию генов на трёх уровнях: они генерируют ключевые метаболиты, непосредственно стирают химические метки на ДНК, РНК и гистонах и действуют как каркасы, реорганизующие хроматиновый аппарат. В совокупности эти функции превращают режим использования топлива клеткой в долговечные инструкции о том, какие гены экспрессировать, помогая сохранять стемнесс или продвигать дифференцировку. Эта концепция объясняет, почему метаболические мутации могут перепрограммировать клетки в сторону ракоподобных состояний и почему изменение питания или метаболических ферментов может модифицировать поведение стволовых клеток. По мере того как исследователи детализируют эти связи, оксидоредуктазы становятся привлекательными мишенями для регенеративной терапии и противораковой стратегии, направленной на перестройку эпигенетического ландшафта путём перенастройки энергетического метаболизма клеток.
Цитирование: Lee, HT., Roe, JS. & Youn, HD. Metabolic oxidoreductases: central regulators of the epigenetic landscapes in stemness. Exp Mol Med 58, 1017–1037 (2026). https://doi.org/10.1038/s12276-026-01687-2
Ключевые слова: метаболизм стволовых клеток, эпигенетическая регуляция, ферменты-оксидоредуктазы, ремоделирование хроматина, раковые стволовые клетки