Clear Sky Science · tr
Hormon yanıt faktörlerinin proteolizine yönelik işlevsel gereksinimlerin çeşitlenmesi
Bitkiler Büyüme Hormonlarını Nasıl Kontrol Altında Tutuyor
Bitkiler, kök ve yapraklardan küçük üreme yapılarının oluşumuna kadar her şeyi şekillendirmek için auxin adlı basit bir hormona dayanır. Ancak basit bir sinyali doğadaki zengin bitki formlarına dönüştürmek sıkı bir kontrol gerektirir. Bu çalışma, ciğerotu bitkilerinin bazı ana düzenleyici proteinleri seçici olarak yok ederek auxine duyarlılıklarını nasıl ince ayar yaptığını araştırıyor; neden bazı proteinlerin hızla parçalanması gerektiğini, bazılarının ise güvenle kalabileceğini ortaya koyuyor.
Auxin Yanıtlarının Arkasındaki İtiş–Çekiş Ekibi
Auxin, bitki hücrelerinin çekirdeğinde çalışan bir yol aracılığıyla genleri açıp kapayarak etkisini gösterir. Bu yolun merkezinde, DNA’ya bağlanıp hormonun hangi genleri etkileyeceğini kontrol eden Auxin Yanıt Faktörleri (ARF'ler) bulunur. Aynı DNA bölgeleri için rekabet eden iki ana ARF türü vardır: A sınıfı ARF’ler genellikle auxin karşısında genleri aktive ederken, B sınıfı ARF’ler genleri kapatır ve sistemde fren görevi görür. Her hücrede bu aktive ediciler ile baskılayıcılar arasındaki denge, o hücrenin auxine ne kadar güçlü yanıt vereceğini ve dolayısıyla bitkinin nasıl gelişeceğini belirler.
Yıkım Tasarım Özelliğine Ne Zaman Dönüşür
Yakın zamandaki çalışmalar, birçok ARF’nin doğuştan kararsız olduğunu gösterdi: DNA bağlanma ucunda taşımış oldukları yerleşik bir işaret onları proteazom adlı hücrenin protein parçalayıcı mekanizması tarafından yıkılmaya hedefler. Ciğerotu Marchantia polymorpha’da her ARF sınıfı tek bir genle temsil edildiği için bu süreci çalışmak daha sade bir sistem sağlar. Yazarlar, B sınıfı ARF olan MpARF2’nin çürüme motifi içindeki tek tek amino asitleri sistematik olarak değiştirdiler. MpARF2’yi kararsız yapan unsurun tek bir pozitif yüklü yapı taşı, bir arginin olduğunu keşfettiler. Pozitif yükün kaldırılması veya yakındaki yüklerin tersine çevrilmesi proteini güçlü şekilde stabilize ederken, aynı bölgedeki birçok diğer değişiklikin ise az etkisi oldu. A sınıfı aktive edici MpARF1’deki karşılık gelen bir arginin benzer bir biyokimyasal rol oynadığı görüldü; bu da bu çürüme sinyalinin ortak eski bir kökene işaret ediyor.
Neden Bir Protein Hızla Kaybolmalı Diğeri Uzun Süre Kalabilir
Her iki MpARF1 ve MpARF2 bu korunan motif aracılığıyla yıkılabilse de, yıkıma olan biyolojik bağımlılıkları çarpıcı biçimde farklıdır. Araştırmacılar MpARF1’i stabilize ettiklerinde, aktive edicinin seviyeleri ılımlı şekilde yükseldi ancak bitki büyümesi, auxin duyarlılığı ve gen ifade desenleri büyük ölçüde normal kaldı. Buna karşılık MpARF2’yi stabilize etmenin dramatik sonuçları oldu. Araştırma ekibi floresan işaretleyiciler ve proteazom inhibitörleri kullanarak MpARF2’nin bitkinin yaşam döngüsü boyunca parçalanmasını haritaladı. Normal bitkilerde MpARF2’nin birçok dokuda üretildiğini ama özellikle büyüme uçları ve rizoid oluşturan hücrelerde hızla ortadan kaldırıldığını buldular. Yıkım engellendiğinde MpARF2 neredeyse her yerde birikti ve bitkiler düz thalli, hava gözenekleri, gemma kupaları ve üreme yapılarını oluşturmakta şiddetli bozukluklar gösterdi.
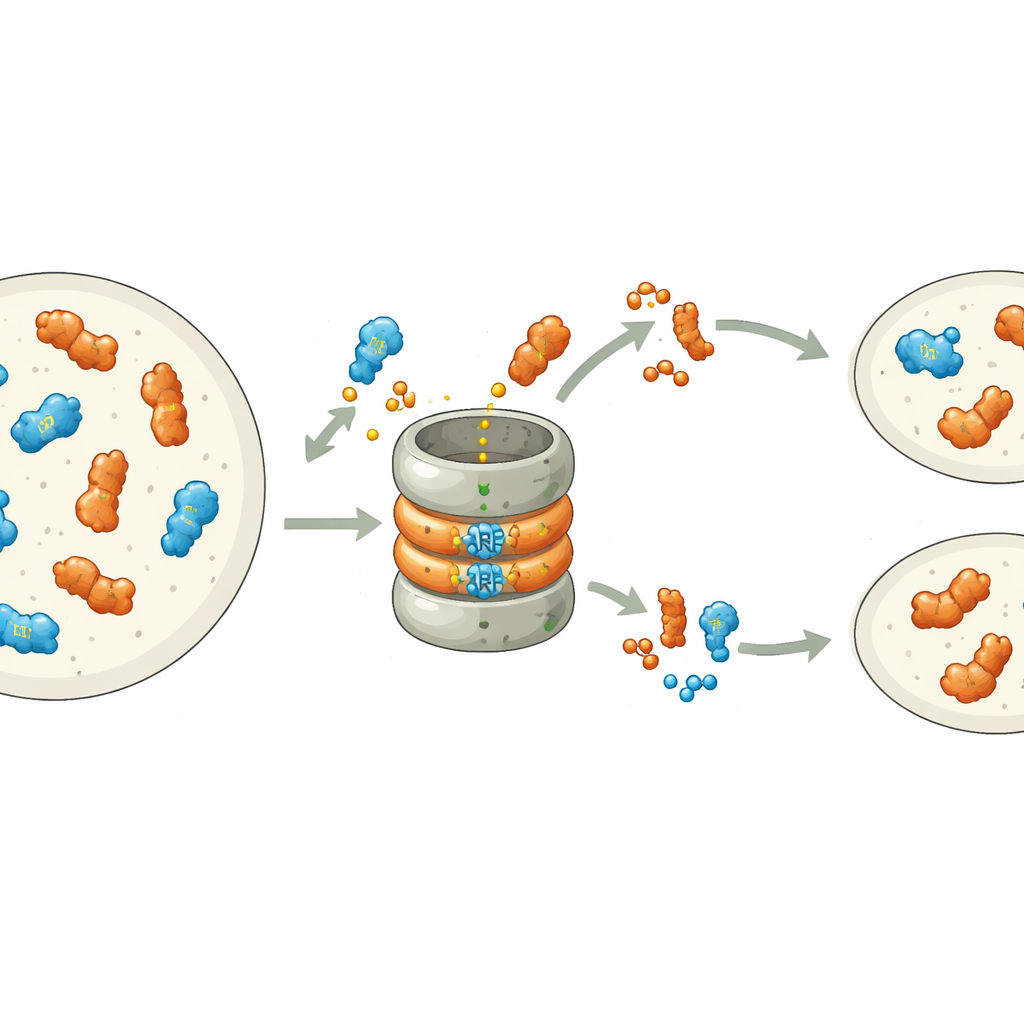
Moleküler Bir Frenin Kontrolündeki Yaşam Döngüsü
Protein kararlılığını doğrudan işleve bağlamak için yazarlar genetik mutantları, akıllıca tasarlanmış ve anahtarlanabilir MpARF2 varyantlarıyla birleştirdiler. Seçilen zamanlarda yıkıma dirençli MpARF2’yi açmak, eklenen auxine bitkilerin yanıtını keskin şekilde düşürdü ve gemma kaplarının ile erkek üreme saplarının normal gelişimini engelledi. Güçlü işlev kaybı mutantını normal bir MpARF2 ile tamamlamak büyüme ve auxin’e duyarlı gen aktivitesini kısmen geri getirirken, stabilize edilmiş bir versiyon bunu başaramadı. Tüm genom düzeyinde RNA dizilemesi, MpARF2 yıkılamadığında tipik auxin’e yanıt veren genlerin tepki vermediğini, oysa ilgisiz pek çok genin düzensiz düzenlendiğini doğruladı. Kısacası, MpARF2 yalnızca hücrenin proteoliz yoluyla seviyesini sürekli budamasına izin verdiği zaman doğru rolünü yerine getiriyor.
Bitki Çeşitliliği ve Kontrolü İçin Anlamı
Çalışma, antik bir yıkım sinyalini paylaşmalarına rağmen A sınıfı ve B sınıfı ARF’lerin artık yok edilmeye çok farklı biçimlerde bağımlı olduklarını ortaya koyuyor. Aktivasyon görevindeki MpARF1 yıkımın azalmasını az etkiyle tolere edebilir; bu muhtemelen ayrı baskılayıcı proteinler gibi diğer güvenlik mekanizmalarının aktivitesini kontrol altında tutmasındandır. Ancak baskılayıcı MpARF2, auxin sistemini ezmemek için sürekli olarak kaldırılmak zorundadır. Bu ayrışma muhtemelen bitkiler karaya çıktıkça evrimleşti ve basit bir hormonun dokuya özgü ve ince ayarlı gelişim programlarını sürüklemesine izin verdi. Seçici protein yıkımının hormon yanıtlarını nasıl şekillendirdiğini göstererek çalışma, bitkilerin ortak bir kimyasal sinyali birçok farklı forma nasıl çevirdiğini açıklamaya yardımcı oluyor ve büyümeyi doğrudan hormon yerine moleküler frenlere hedefleyerek manipüle etmenin yeni yollarını öneriyor.
Atıf: de Roij, M., Heijdra, E., Lamers, J. et al. Diversification of functional requirements for proteolysis of auxin response factors. Nat Commun 17, 3917 (2026). https://doi.org/10.1038/s41467-026-70440-0
Anahtar kelimeler: auxin sinyalleşmesi, protein yıkımı, Auxin Yanıt Faktörleri, Marchantia polymorpha, bitki gelişimi