Clear Sky Science · pt
Diversificação dos requisitos funcionais para a proteólise de fatores de resposta a auxina
Como as plantas mantêm seu hormônio de crescimento sob controle
As plantas dependem de um químico simples, o hormônio auxina, para moldar desde raízes e folhas até pequenas estruturas reprodutivas. Mas transformar um sinal simples na rica variedade de formas vegetais que vemos na natureza exige controle rigoroso. Este estudo explora como plantas hepáticas ajustam sua sensibilidade à auxina ao destruir seletivamente proteínas regulatórias-chave, revelando por que algumas dessas proteínas precisam ser rapidamente degradadas enquanto outras podem permanecer sem perigo.
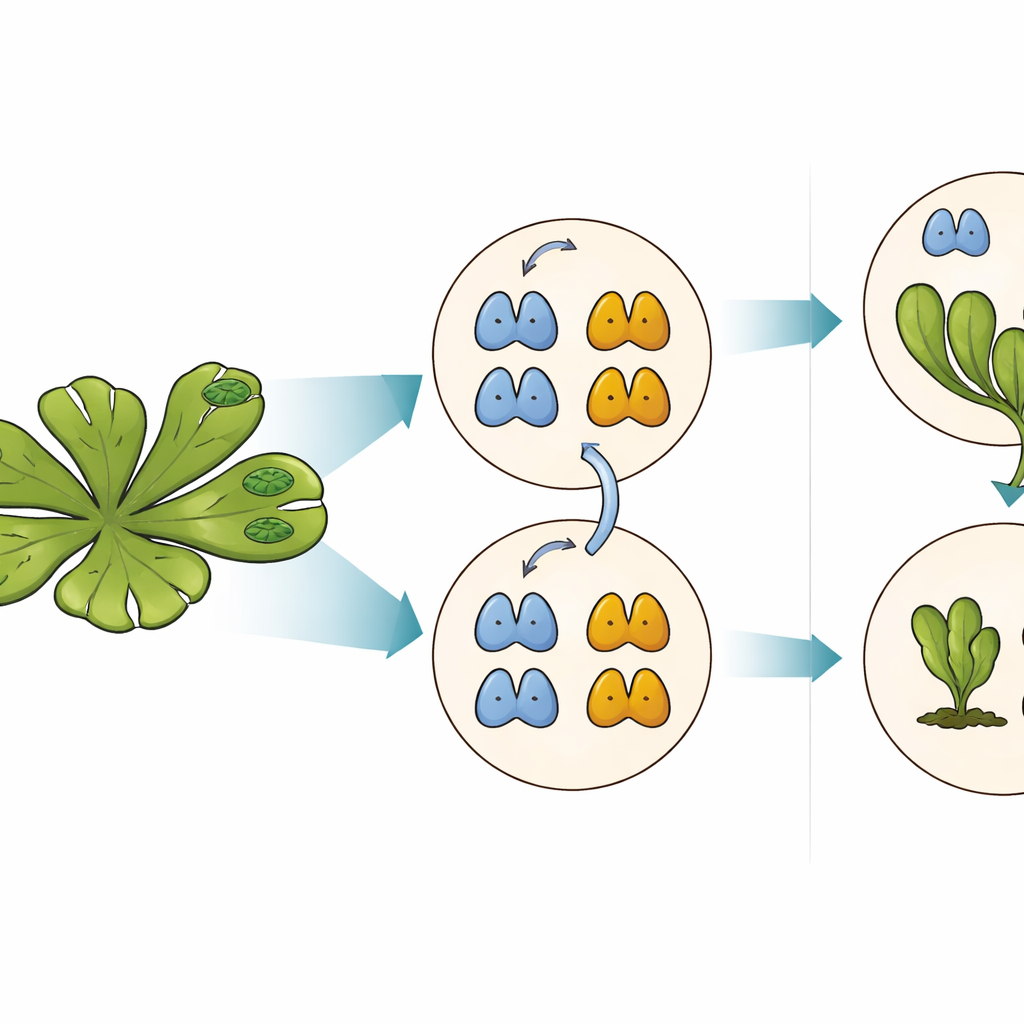
A equipe de empurrar–puxar por trás das respostas à auxina
A auxina age principalmente ligando e desligando genes por meio de uma via nuclear dentro das células vegetais. No centro dessa via estão os Fatores de Resposta à Auxina, ou ARFs, que se ligam ao DNA e controlam quais genes respondem ao hormônio. Dois tipos principais de ARFs competem pelos mesmos sítios no DNA: os ARFs da classe A geralmente ativam genes em resposta à auxina, enquanto os ARFs da classe B reprimem genes e atuam como freios no sistema. O equilíbrio entre esses ativadores e repressores em cada célula determina quão fortemente essa célula reage à auxina e, por sua vez, como a planta se desenvolve.
Quando a destruição se torna um recurso de projeto
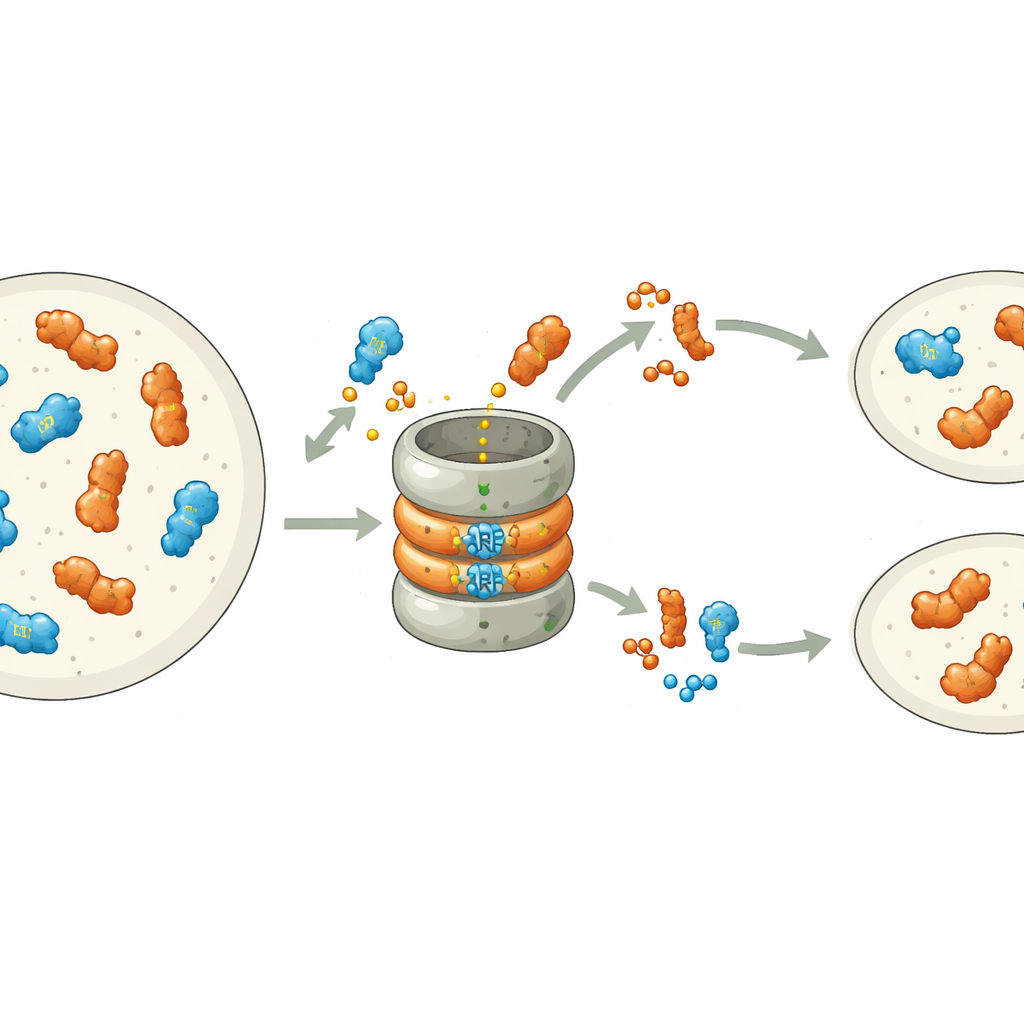
Trabalhos recentes mostraram que muitos ARFs são intrinsecamente instáveis: eles carregam um sinal embutido em sua ponta de ligação ao DNA que os marca para destruição pela maquinaria de trituração de proteínas da célula, o proteassoma. Em Marchantia polymorpha, cada classe de ARF é representada por um único gene, tornando o sistema ideal para estudar esse processo. Os autores modificaram sistematicamente aminoácidos individuais no motivo de degradação do ARF da classe B, MpARF2. Descobriram que um único bloco construtor carregado positivamente, uma arginina, é crítico para tornar MpARF2 instável. Remover sua carga positiva ou inverter cargas próximas estabiliza fortemente a proteína, enquanto muitas outras alterações na mesma região têm pouco efeito. Uma arginina correspondente no ativador da classe A, MpARF1, desempenha um papel bioquímico semelhante, apontando para uma origem antiga compartilhada desse sinal de degradação.
Por que uma proteína deve desaparecer rapidamente e a outra pode permanecer
Embora tanto MpARF1 quanto MpARF2 possam ser degradados por meio desse motivo conservado, sua dependência biológica da degradação é marcadamente distinta. Quando os pesquisadores estabilizaram MpARF1, o ativador, os níveis proteicos aumentaram modestamente, mas o crescimento da planta, a sensibilidade à auxina e os padrões de expressão gênica permaneceram em grande parte normais. Em contraste, estabilizar MpARF2, o repressor, teve consequências dramáticas. Usando marcadores fluorescentes e inibidores do proteassoma, a equipe mapeou a degradação de MpARF2 ao longo do ciclo de vida da planta. Eles descobriram que, em plantas normais, MpARF2 é produzido em muitos tecidos, mas removido rapidamente, especialmente em ápices em crescimento e células formadoras de rizoides. Quando a degradação foi bloqueada, MpARF2 se acumulou quase em todos os lugares, e as plantas apresentaram defeitos severos na formação de talos achatados, poros de ar, cálices de gema (gemma cups) e estruturas reprodutivas.
Ciclo de vida sob controle de um freio molecular
Para conectar diretamente a estabilidade proteica à função, os autores combinaram mutantes genéticos com versões engenhosas e comutáveis de MpARF2. Ativar uma versão resistente à degradação de MpARF2 em tempos escolhidos reduziu fortemente a resposta das plantas à auxina adicionada e impediu o desenvolvimento normal de cálices de gema e de hastes reprodutivas masculinas. Complementar um mutante de forte perda de função com um MpARF2 normal restaurou parcialmente o crescimento e a atividade gênica responsiva à auxina, enquanto uma versão estabilizada não o fez. Sequenciamento de RNA em todo o genoma confirmou que, quando MpARF2 não pode ser degradado, genes típicos responsivos à auxina deixam de reagir, ao passo que muitos genes não relacionados ficam mal regulados. Em resumo, MpARF2 só desempenha sua função adequada quando a célula pode continuamente ajustar seus níveis por meio da proteólise.
O que isso significa para a diversidade e o controle das plantas
O trabalho revela que, apesar de compartilharem um sinal de degradação ancestral, ARFs das classes A e B agora dependem de maneira muito diferente de serem destruídos. O ativador MpARF1 pode tolerar redução na degradação com pouco impacto, possivelmente porque outras salvaguardas, como proteínas repressoras separadas, mantêm sua atividade sob controle. O repressor MpARF2, entretanto, deve ser continuamente removido para evitar sobrecarregar o sistema de auxina. Essa divergência provavelmente evoluiu quando as plantas colonizaram a terra, permitindo que um hormônio simples dirigisse programas de desenvolvimento finamente ajustados e específicos por tecido. Ao mostrar como a destruição seletiva de proteínas esculpe respostas hormonais, o estudo ajuda a explicar como as plantas traduzem um sinal químico comum em muitas formas distintas e sugere novas maneiras de manipular o crescimento visando os freios moleculares em vez do próprio hormônio.
Citação: de Roij, M., Heijdra, E., Lamers, J. et al. Diversification of functional requirements for proteolysis of auxin response factors. Nat Commun 17, 3917 (2026). https://doi.org/10.1038/s41467-026-70440-0
Palavras-chave: sinalização por auxina, degradação de proteínas, Fatores de Resposta à Auxina, Marchantia polymorpha, desenvolvimento de plantas