Clear Sky Science · es
Diversificación de los requisitos funcionales para la proteólisis de los factores de respuesta a auxina
Cómo las plantas mantienen controlada su hormona de crecimiento
Las plantas dependen de una molécula sencilla, la hormona auxina, para moldear desde raíces y hojas hasta pequeñas estructuras reproductivas. Pero convertir una señal simple en la rica variedad de formas vegetales que observamos en la naturaleza requiere un control estricto. Este estudio explora cómo los hepáticas afinan su sensibilidad a la auxina mediante la destrucción selectiva de proteínas reguladoras clave, revelando por qué algunas de estas proteínas deben degradarse rápidamente mientras que otras pueden permanecer sin problema.
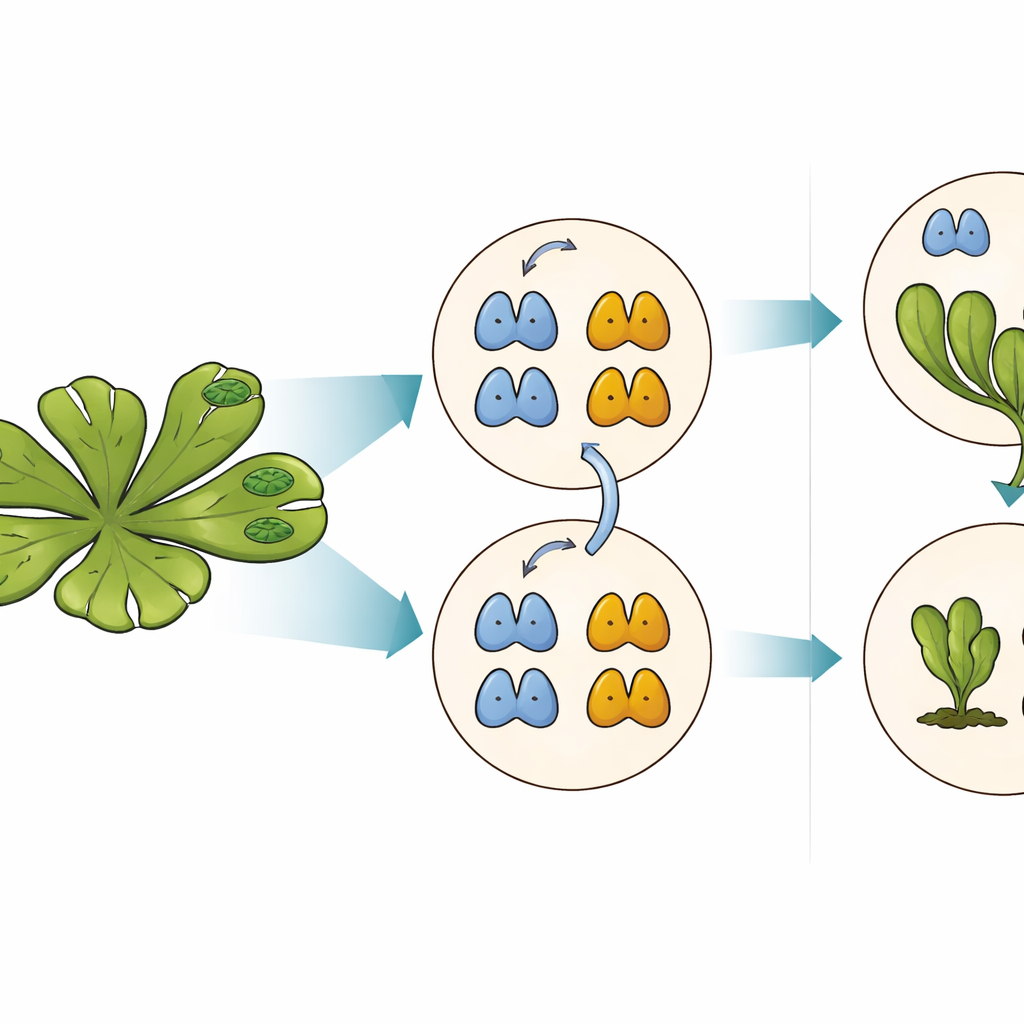
El equipo de empuje y freno detrás de las respuestas a la auxina
La auxina actúa principalmente activando o desactivando genes a través de una vía nuclear dentro de las células vegetales. En el centro de esta vía están los Factores de Respuesta a Auxina, o ARF, que se unen al ADN y controlan qué genes responden a la hormona. Dos tipos principales de ARF compiten por los mismos sitios del ADN: los ARF de clase A suelen activar genes en respuesta a la auxina, mientras que los ARF de clase B reprimen genes y actúan como frenos del sistema. El equilibrio entre estos activadores y represores en cada célula determina cuán intensamente esa célula reacciona a la auxina y, a su vez, cómo se desarrolla la planta.
Cuando la destrucción se convierte en una característica de diseño
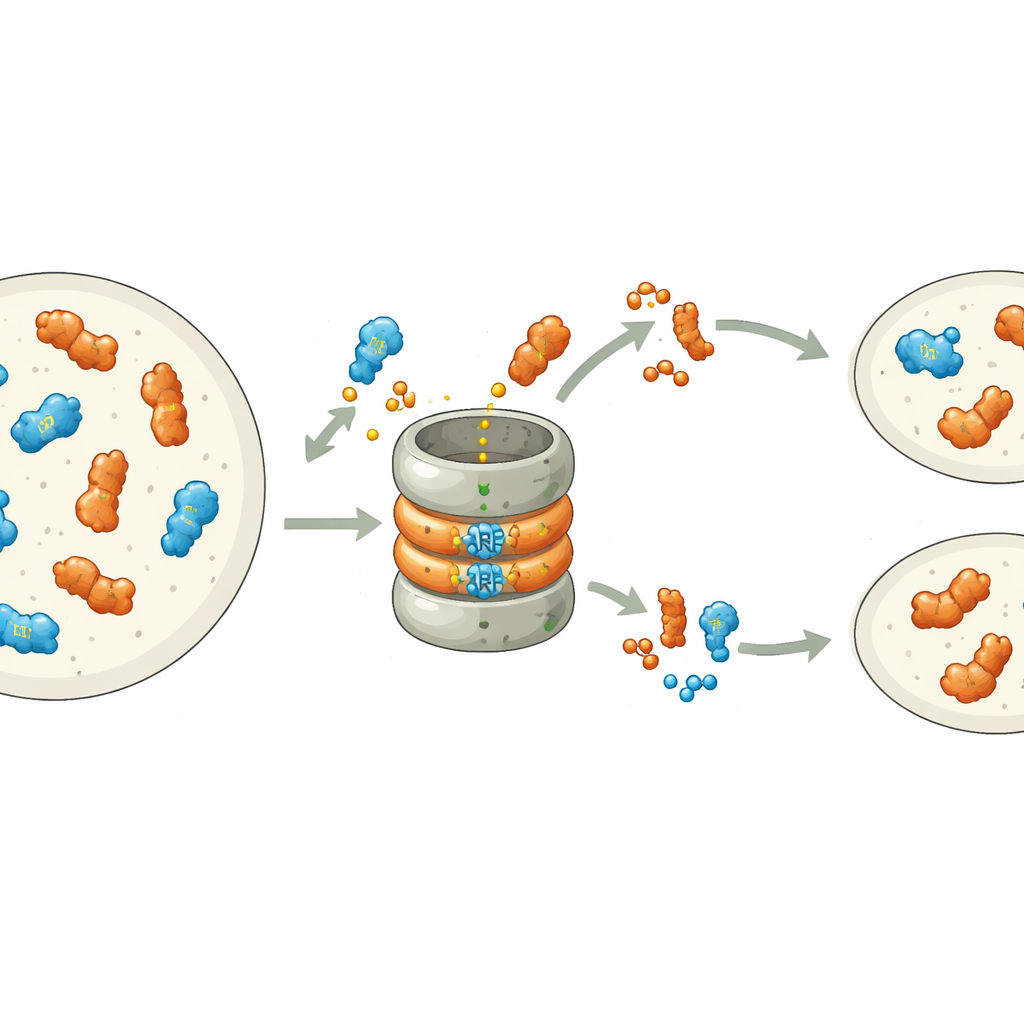
Trabajos recientes mostraron que muchos ARF son inherentemente inestables: llevan una señal integrada en su extremo de unión al ADN que los marca para la destrucción por la maquinaria celular de degradación de proteínas, el proteasoma. En la hepática Marchantia polymorpha, cada clase de ARF está representada por un único gen, lo que la convierte en un sistema claro para estudiar este proceso. Los autores cambiaron sistemáticamente aminoácidos individuales en el motivo de degradación del ARF de clase B, MpARF2. Descubrieron que un único bloque positivo, una arginina, es crítico para que MpARF2 sea inestable. Eliminar su carga positiva o invertir cargas cercanas estabiliza fuertemente la proteína, mientras que muchos otros cambios en la misma región tienen poco efecto. Una arginina correspondiente en el activador de clase A, MpARF1, desempeña un papel bioquímico similar, lo que apunta a un origen antiguo compartido de esta señal de degradación.
Por qué una proteína debe desaparecer rápidamente y la otra puede persistir
Aunque tanto MpARF1 como MpARF2 pueden ser degradados mediante este motivo conservado, su dependencia biológica de la degradación difiere notablemente. Cuando los investigadores estabilizaron MpARF1, el activador, los niveles de proteína aumentaron modestamente, pero el crecimiento de la planta, la sensibilidad a la auxina y los patrones de expresión génica siguieron siendo en gran medida normales. En contraste, estabilizar MpARF2, el represor, tuvo consecuencias dramáticas. Usando etiquetas fluorescentes e inhibidores del proteasoma, el equipo cartografió la degradación de MpARF2 a lo largo del ciclo de vida de la planta. Encontraron que en plantas normales MpARF2 se produce en muchos tejidos pero se elimina rápidamente, especialmente en puntas en crecimiento y células formadoras de rizoides. Cuando se bloqueó la degradación, MpARF2 se acumuló casi por todas partes y las plantas mostraron defectos severos en la formación de talos planos, poros aéreos, copas de gemas y estructuras reproductivas.
Ciclo de vida bajo control de un freno molecular
Para conectar la estabilidad proteica directamente con la función, los autores combinaron mutantes genéticos con versiones ingenierizadas y conmutables de MpARF2. Activar una versión de MpARF2 resistente a la degradación en tiempos elegidos redujo drásticamente la respuesta de las plantas a la auxina añadida e impidió el desarrollo normal de las copas de gemas y de los pedúnculos reproductivos masculinos. Complementar un mutante de pérdida de función intensa con un MpARF2 normal restauró parcialmente el crecimiento y la actividad de genes responsivos a la auxina, mientras que una versión estabilizada no lo hizo. La secuenciación de ARN a nivel genómico confirmó que cuando MpARF2 no puede degradarse, los genes típicamente responsivos a la auxina dejan de reaccionar, mientras que muchos genes no relacionados quedan desregulados. En resumen, MpARF2 solo desempeña su papel correcto cuando la célula puede recortar continuamente sus niveles mediante proteólisis.
Lo que esto significa para la diversidad y el control vegetal
El trabajo revela que, a pesar de compartir una señal de degradación antigua, los ARF de clase A y B ahora dependen de manera muy diferente de ser destruidos. El activador MpARF1 puede tolerar una reducción en su degradación con poco impacto, posiblemente porque otras salvaguardas, como proteínas represoras separadas, mantienen su actividad bajo control. El represor MpARF2, sin embargo, debe eliminarse continuamente para evitar que domine el sistema de auxina. Esta divergencia probablemente evolucionó cuando las plantas colonizaron la tierra, permitiendo que una hormona simple impulse programas de desarrollo finamente ajustados y específicos por tejido. Al mostrar cómo la destrucción selectiva de proteínas esculpe las respuestas hormonales, el estudio ayuda a explicar cómo las plantas traducen una señal química común en muchas formas distintas y sugiere nuevas maneras de manipular el crecimiento apuntando a los frenos moleculares en lugar de a la hormona misma.
Cita: de Roij, M., Heijdra, E., Lamers, J. et al. Diversification of functional requirements for proteolysis of auxin response factors. Nat Commun 17, 3917 (2026). https://doi.org/10.1038/s41467-026-70440-0
Palabras clave: señalización por auxina, degradación de proteínas, Factores de Respuesta a Auxina, Marchantia polymorpha, desarrollo vegetal