Clear Sky Science · fr
Diversification des exigences fonctionnelles pour la protéolyse des facteurs de réponse à l'auxine
Comment les plantes contrôlent leur hormone de croissance
Les plantes s'appuient sur une molécule simple, l'hormone auxine, pour façonner tout, des racines et des feuilles aux petites structures reproductrices. Mais transformer un signal simple en la riche variété de formes végétales observées dans la nature exige un contrôle strict. Cette étude examine comment les hépatiques adaptent leur sensibilité à l'auxine en détruisant sélectivement des protéines régulatrices clés, révélant pourquoi certaines de ces protéines doivent être rapidement éliminées tandis que d'autres peuvent persister sans danger.
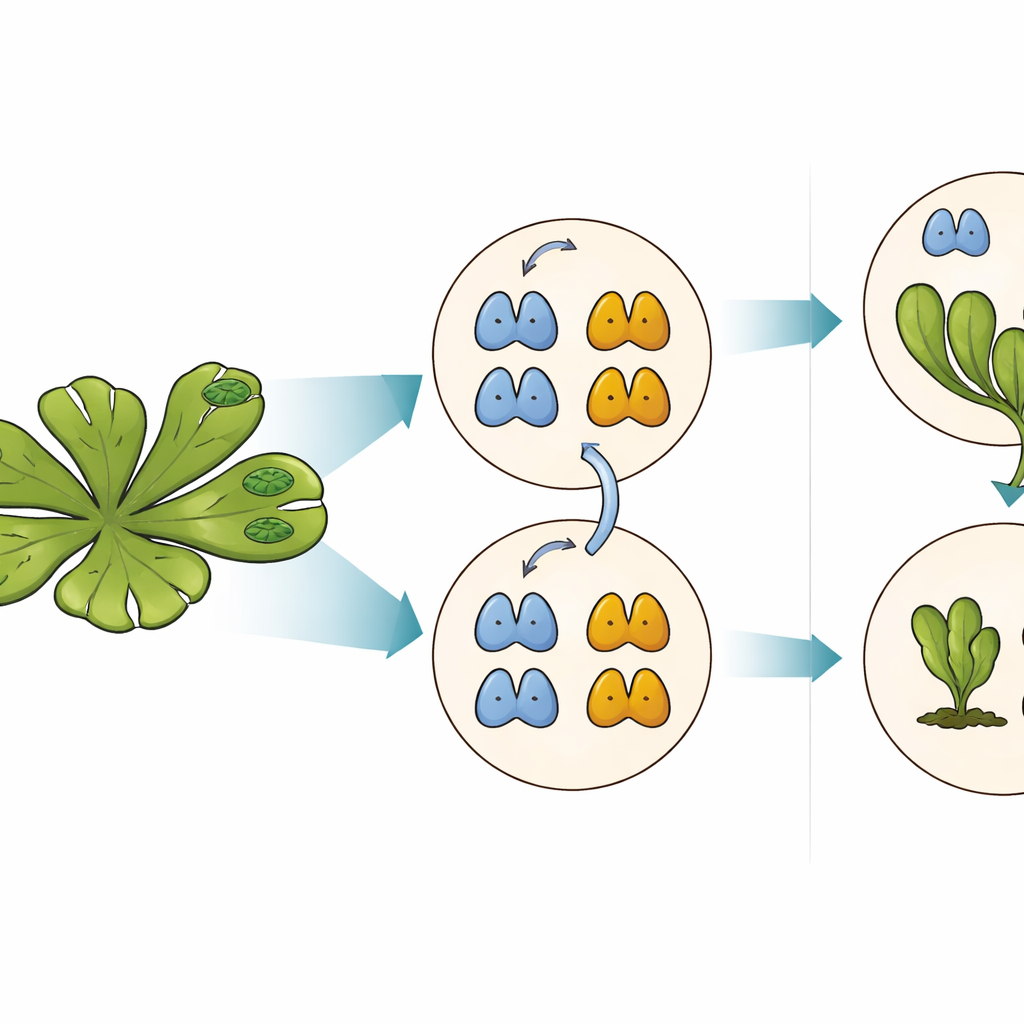
L'équipe antagoniste derrière les réponses à l'auxine
L'auxine agit principalement en activant ou en réprimant des gènes via une voie nucléaire à l'intérieur des cellules végétales. Au cœur de cette voie se trouvent les facteurs de réponse à l'auxine, ou ARF, qui se lient à l'ADN et contrôlent quels gènes répondent à l'hormone. Deux types principaux d'ARF se disputent les mêmes sites d'ADN : les ARF de classe A activent généralement les gènes en réponse à l'auxine, tandis que les ARF de classe B éteignent les gènes et jouent le rôle de freins sur le système. L'équilibre entre ces activateurs et répresseurs dans chaque cellule détermine l'intensité de la réponse cellulaire à l'auxine et, par conséquent, le développement de la plante.
Quand la destruction devient un élément de conception
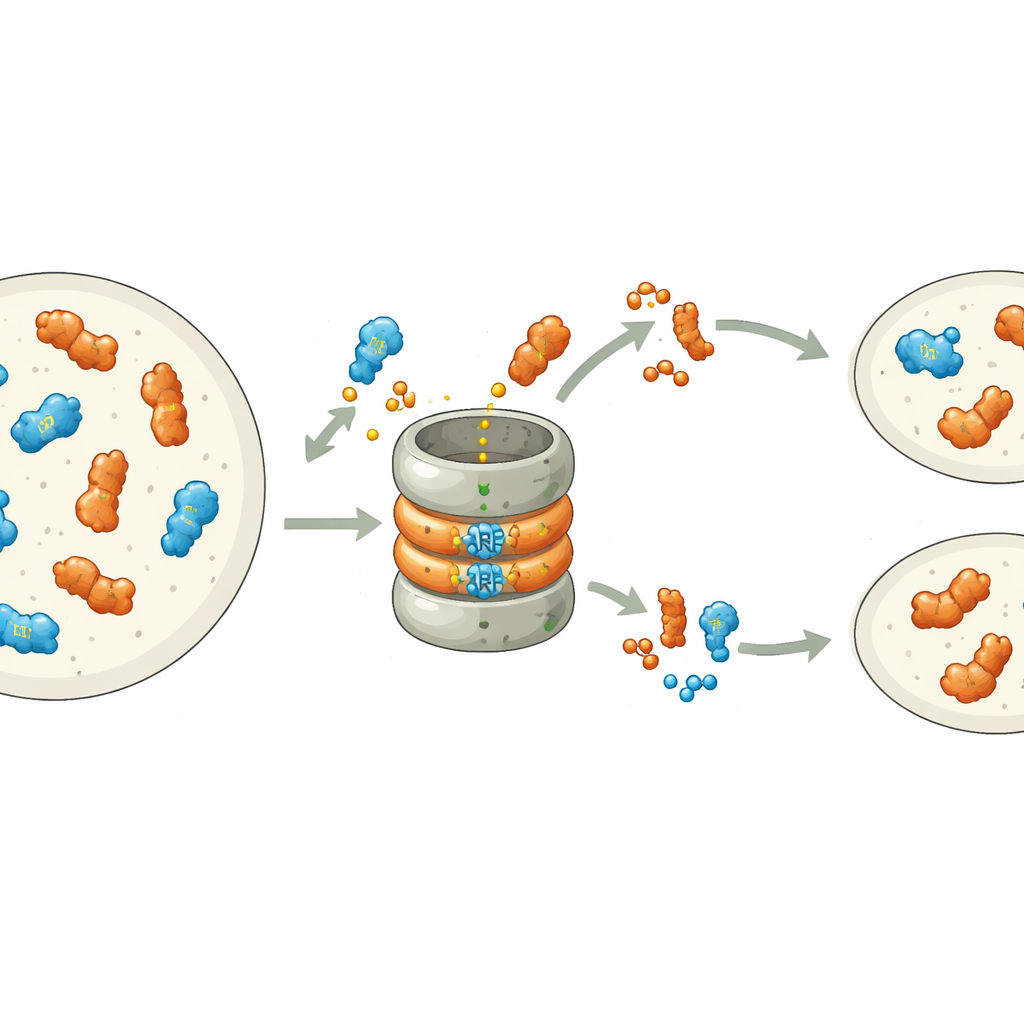
Des travaux récents ont montré que de nombreux ARF sont intrinsèquement instables : ils portent un signal intégré à leur extrémité de liaison à l'ADN qui les marque pour destruction par la machinerie de déchiquetage des protéines de la cellule, le protéasome. Chez la hépatiques Marchantia polymorpha, chaque classe d'ARF est représentée par un seul gène, ce qui en fait un système simple pour étudier ce processus. Les auteurs ont modifié systématiquement des acides aminés individuels du motif de dégradation de l'ARF de classe B, MpARF2. Ils ont découvert qu'un seul élément chargé positivement, une arginine, est crucial pour rendre MpARF2 instable. Supprimer sa charge positive ou inverser les charges à proximité stabilise fortement la protéine, tandis que de nombreux autres changements dans la même région ont peu d'effet. Une arginine correspondante dans l'activatrice de classe A, MpARF1, joue un rôle biochimique similaire, suggérant une origine ancienne partagée de ce signal de dégradation.
Pourquoi une protéine doit disparaître vite et l'autre peut persister
Bien que MpARF1 et MpARF2 puissent tous deux être dégradés via ce motif conservé, leur dépendance biologique à la dégradation diffère fortement. Lorsque les chercheurs ont stabilisé MpARF1, l'activateur, le niveau de protéine a augmenté modestement, mais la croissance des plantes, la sensibilité à l'auxine et les profils d'expression génique sont restés globalement normaux. En revanche, la stabilisation de MpARF2, le répresseur, a eu des conséquences spectaculaires. En utilisant des marqueurs fluorescents et des inhibiteurs du protéasome, l'équipe a cartographié la dégradation de MpARF2 tout au long du cycle de vie de la plante. Ils ont constaté que dans les plantes normales MpARF2 est produit dans de nombreux tissus mais rapidement éliminé, en particulier dans les pointes de croissance et les cellules formant les rhizoïdes. Lorsque la dégradation était bloquée, MpARF2 s'accumulait presque partout et les plantes présentaient de sévères défauts de formation des thalles plats, des pores aériens, des coupes de gemma et des structures reproductrices.
Cycle de vie contrôlé par un frein moléculaire
Pour lier directement la stabilité des protéines à leur fonction, les auteurs ont combiné des mutants génétiques avec des versions ingénieuses et commutables de MpARF2. Activer un MpARF2 résistant à la dégradation à des moments choisis réduisait fortement la réponse des plantes à l'auxine ajoutée et empêchait le développement normal des coupes de gemma et des hampe reproductrices mâles. Compenser un mutant à forte perte de fonction par un MpARF2 normal rétablit partiellement la croissance et l'activité des gènes réactifs à l'auxine, tandis qu'une version stabilisée ne le faisait pas. Le séquençage de l'ARN à l'échelle du génome a confirmé que lorsque MpARF2 ne peut pas être dégradé, les gènes normalement réactifs à l'auxine ne répondent pas, tandis que de nombreux gènes non liés sont mal régulés. En bref, MpARF2 n'accomplit son rôle correct que lorsque la cellule peut en réduire continuellement le niveau par protéolyse.
Ce que cela signifie pour la diversité et le contrôle des plantes
Ce travail révèle que, malgré le partage d'un signal de dégradation ancien, les ARF de classe A et de classe B dépendent aujourd'hui très différemment de leur destruction. L'activateur MpARF1 peut tolérer une réduction de sa dégradation sans grand impact, peut‑être parce que d'autres garde‑fous, comme des protéines répressives distinctes, limitent son activité. Le répresseur MpARF2, en revanche, doit être continuellement éliminé pour ne pas submerger le système auxine. Cette divergence a probablement évolué lors de la colonisation des terres par les plantes, permettant à une hormone simple de piloter des programmes de développement finement réglés et spécifiques aux tissus. En montrant comment la destruction sélective des protéines sculpte les réponses hormonales, l'étude aide à expliquer comment les plantes traduisent un signal chimique commun en de nombreuses formes distinctes et suggère de nouvelles façons de manipuler la croissance en visant les freins moléculaires plutôt que l'hormone elle‑même.
Citation: de Roij, M., Heijdra, E., Lamers, J. et al. Diversification of functional requirements for proteolysis of auxin response factors. Nat Commun 17, 3917 (2026). https://doi.org/10.1038/s41467-026-70440-0
Mots-clés: signalisation auxine, dégradation des protéines, Facteurs de réponse à l'auxine, Marchantia polymorpha, développement des plantes