Clear Sky Science · ru
Диверсификация функциональных требований к протеолизу факторов ответа на ауксин
Как растения держат свой гормон роста под контролем
Растения полагаются на простой химический сигнал — гормон ауксин, который формирует всё: от корней и листьев до крошечных репродуктивных структур. Но превращение простого сигнала в богатое разнообразие растительных форм требует строгого контроля. В этом исследовании рассматривается, как печёночные мхи точно настраивают чувствительность к ауксину посредством выборочного разрушения ключевых регуляторных белков, что объясняет, почему некоторые из этих белков должны быстро расщепляться, тогда как другие могут безопасно оставаться в клетке.
Команда «толчок–тормоз» в основе ответов на ауксин
Ауксин действует в основном через включение или выключение генов посредством ядерного пути внутри растительных клеток. В центре этого пути находятся факторы ответа на ауксин (ARF), которые связываются с ДНК и регулируют, какие гены реагируют на гормон. Две основные группы ARF соперничают за одни и те же сайты ДНК: ARF класса A обычно активируют гены в ответ на ауксин, тогда как ARF класса B подавляют гены и работают как тормоза системы. Баланс между этими активаторами и репрессорами в каждой клетке определяет, насколько сильно клетка отвечает на ауксин и, следовательно, как развивается растение.
Когда разрушение становится элементом дизайна
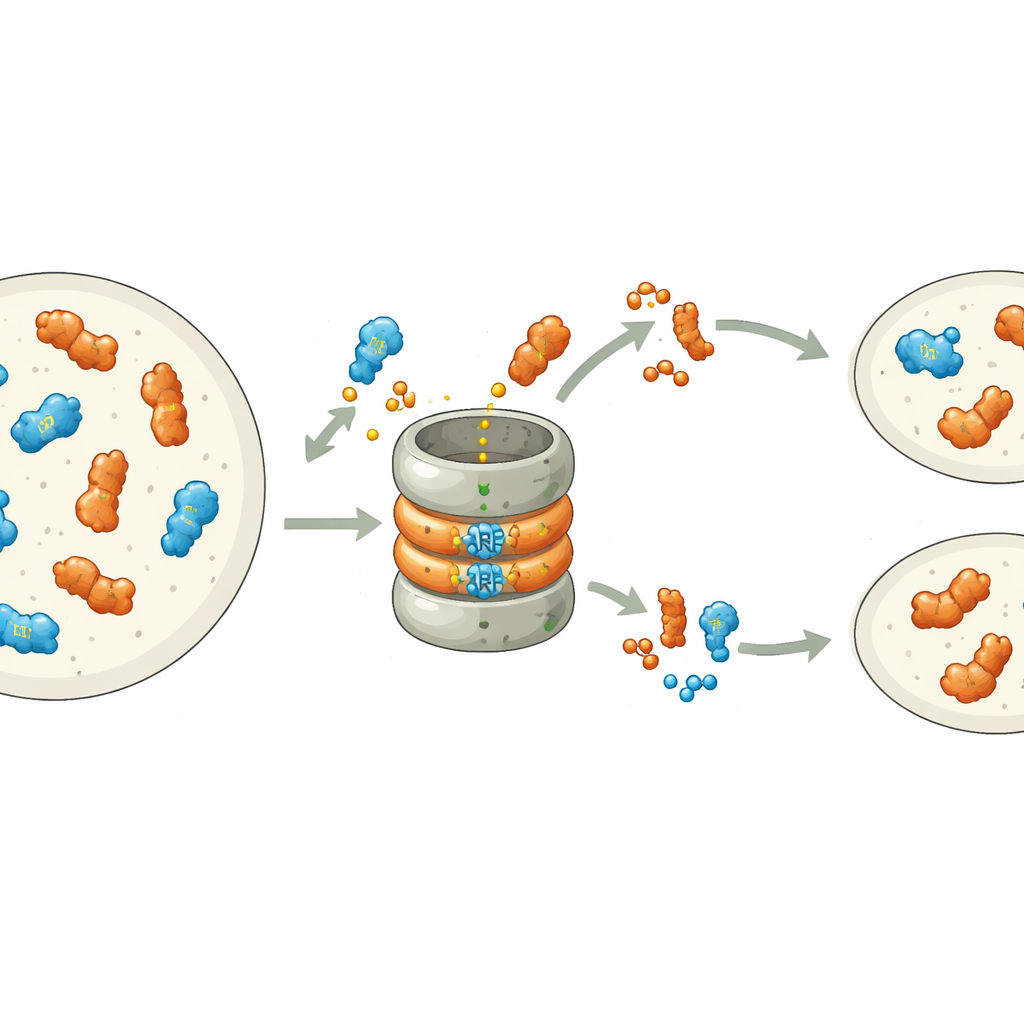
Недавние исследования показали, что многие ARF по своей природе нестабильны: в их концевой домен, связывающий ДНК, встроен сигнал, помечающий их для уничтожения белковым комплексом протеасомы. В печёночном мхе Marchantia polymorpha каждый класс ARF представлен одним геном, что делает эту систему удобной для изучения процесса. Авторы методично изменяли отдельные аминокислоты в мотивах деградации ARF класса B, MpARF2. Они обнаружили, что одна положительно заряженная аминокислота, аргинин, критически важна для нестабильности MpARF2. Устранение положительного заряда или изменение близлежащих зарядов сильно стабилизирует белок, тогда как многие другие изменения в том же регионе мало влияют. Соответствующий аргинин в активаторе класса A, MpARF1, выполняет аналогичную биохимическую роль, указывая на общее древнее происхождение этого сигнала деградации.
Почему один белок должен исчезать быстро, а другой может оставаться дольше
Несмотря на то, что и MpARF1, и MpARF2 могут деградировать через этот консервативный мотив, их биологическая зависимость от деградации резко различается. При стабилизации MpARF1, активатора, уровень белка немного повышается, но рост растения, чувствительность к ауксину и профиль экспрессии генов в целом остаются нормальными. Напротив, стабилизация MpARF2, репрессора, приводит к драматическим последствиям. С помощью флуоресцентных меток и ингибиторов протеасомы исследователи проследили распад MpARF2 на протяжении жизненного цикла растения. Они выяснили, что у нормальных растений MpARF2 синтезируется во многих тканях, но быстро удаляется, особенно в растущих кончиках и клетках, формирующих ризоиды. При блокировке деградации MpARF2 накапливается почти повсеместно, и растения демонстрируют серьёзные дефекты в формировании плоских талломов, устьиц воздушных пор, чашек гемм и репродуктивных структур.
Жизненный цикл под контролем молекулярного тормоза
Чтобы напрямую связать стабильность белка с его функцией, авторы сочетали генетические мутанты с хитро сконструированными переключаемыми версиями MpARF2. Включение устойчивой к деградации версии MpARF2 в выбранные моменты резко снижало ответ растений на добавленный ауксин и препятствовало нормальному развитию чашек гемм и мужских репродуктивных стеблей. Дополнение сильного нокдауна нормальной MpARF2 частично восстанавливало рост и активность ауксин‑зависимых генов, тогда как стабилизированная версия этого не делала. Ранжирование транскриптома подтвердило, что при неспособности MpARF2 подвергаться деградации типичные ауксин‑реагирующие гены перестают отвечать, тогда как многие несвязанные гены оказываются неправильно регулируемыми. Коротко говоря, MpARF2 исполняет свою роль только тогда, когда клетка может постоянно подрезать его уровни через протеолиз.
Что это значит для разнообразия и контроля в растениях
Работа показывает, что, несмотря на общее древнее сигнальное мотиво деградации, ARF класса A и класса B теперь по‑разному зависят от разрушения. Активатор MpARF1 может терпеть снижение разрушения с незначительными последствиями, возможно потому, что другие предохранители, например отдельные белки‑репрессоры, сдерживают его активность. Репрессор MpARF2, наоборот, должен постоянно удаляться, чтобы не подавлять систему ауксина. Эта дивергенция, вероятно, эволюционировала по мере колонизации суши растениями, позволяя простому гормону управлять тонко настроенными и тканеспецифическими программами развития. Показав, как выборочное разрушение белков формирует ответы на гормоны, исследование помогает понять, как растения переводят общий химический сигнал в множество различных форм, и открывает новые пути для манипуляции ростом путем нацеливания на молекулярные тормоза, а не на сам гормон.
Цитирование: de Roij, M., Heijdra, E., Lamers, J. et al. Diversification of functional requirements for proteolysis of auxin response factors. Nat Commun 17, 3917 (2026). https://doi.org/10.1038/s41467-026-70440-0
Ключевые слова: сигнализация ауксина, разрушение белков, Факторы ответа на ауксин, Marchantia polymorpha, развитие растений