Clear Sky Science · en
Diversification of functional requirements for proteolysis of auxin response factors
How Plants Keep Their Growth Hormone in Check
Plants rely on a simple chemical, the hormone auxin, to shape everything from roots and leaves to tiny reproductive structures. But turning a simple signal into the rich variety of plant forms we see in nature requires tight control. This study explores how liverwort plants fine‑tune their sensitivity to auxin by selectively destroying key regulatory proteins, revealing why some of these proteins must be rapidly broken down while others can safely stick around.
The Push–Pull Team Behind Auxin Responses
Auxin works largely by switching genes on or off through a nuclear pathway inside plant cells. At the heart of this pathway are Auxin Response Factors, or ARFs, which bind DNA and control which genes respond to the hormone. Two main types of ARFs compete for the same DNA sites: A‑class ARFs generally activate genes in response to auxin, while B‑class ARFs shut genes down and act as brakes on the system. The balance between these activators and repressors in each cell determines how strongly that cell reacts to auxin and, in turn, how the plant develops.
When Destruction Becomes a Design Feature
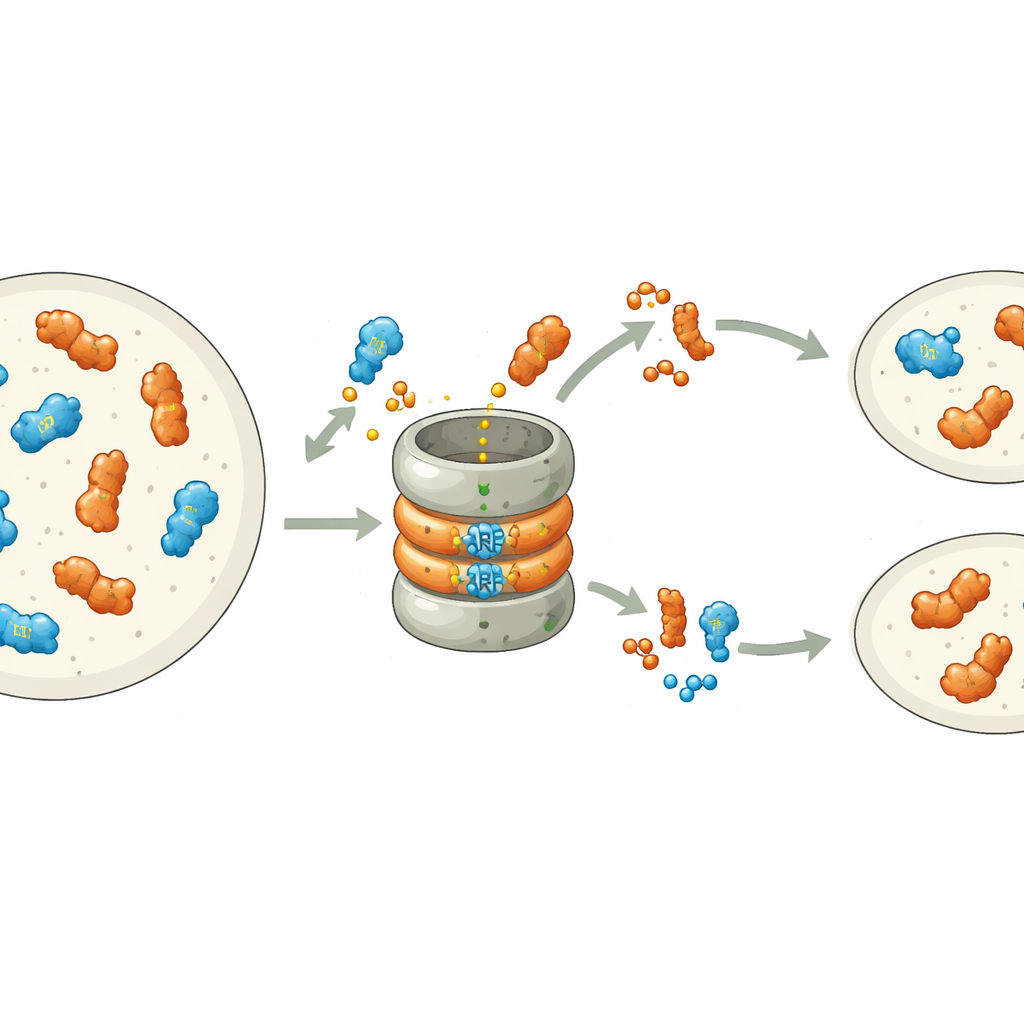
Recent work showed that many ARFs are inherently unstable: they carry a built‑in signal in their DNA‑binding tip that marks them for destruction by the cell’s protein‑shredding machinery, the proteasome. In the liverwort Marchantia polymorpha, each ARF class is represented by a single gene, making it a clean system to study this process. The authors systematically changed individual amino acids in the degradation motif of the B‑class ARF, MpARF2. They discovered that a single positively charged building block, an arginine, is critical for making MpARF2 unstable. Removing its positive charge or flipping nearby charges strongly stabilizes the protein, while many other changes in the same region have little effect. A corresponding arginine in the A‑class activator MpARF1 plays a similar biochemical role, pointing to a shared ancient origin of this degradation signal.
Why One Protein Must Vanish Quickly and the Other Can Linger
Although both MpARF1 and MpARF2 can be degraded through this conserved motif, their biological reliance on degradation is strikingly different. When the researchers stabilized MpARF1, the activator, the protein levels rose modestly, but plant growth, auxin sensitivity, and gene expression patterns remained largely normal. In contrast, stabilizing MpARF2, the repressor, had dramatic consequences. Using fluorescent tags and inhibitors of the proteasome, the team mapped MpARF2 breakdown throughout the plant’s life cycle. They found that in normal plants MpARF2 is made in many tissues but quickly removed, especially in growing tips and rhizoid‑forming cells. When degradation was blocked, MpARF2 accumulated almost everywhere, and plants showed severe defects in forming flat thalli, air pores, gemma cups, and reproductive structures.
Life Cycle Under Control of a Molecular Brake
To connect protein stability directly to function, the authors combined genetic mutants with cleverly engineered, switchable versions of MpARF2. Turning on a degradation‑resistant MpARF2 at chosen times sharply reduced the plants’ response to added auxin and prevented normal development of gemma cups and male reproductive stalks. Complementing a strong loss‑of‑function mutant with a normal MpARF2 partially restored growth and auxin‑responsive gene activity, whereas a stabilized version did not. Genome‑wide RNA sequencing confirmed that when MpARF2 cannot be degraded, typical auxin‑responsive genes fail to react, while many unrelated genes are mis‑regulated. In short, MpARF2 only performs its proper role when the cell can continuously trim its levels through proteolysis.
What This Means for Plant Diversity and Control
The work reveals that, despite sharing an ancient degradation signal, A‑class and B‑class ARFs now depend very differently on being destroyed. The activator MpARF1 can tolerate reduced breakdown with little impact, possibly because other safeguards, such as separate repressor proteins, keep its activity in check. The repressor MpARF2, however, must be continually removed to avoid overwhelming the auxin system. This divergence likely evolved as plants colonized land, allowing a simple hormone to drive finely tuned and tissue‑specific developmental programs. By showing how selective protein destruction sculpts hormone responses, the study helps explain how plants translate a common chemical signal into many distinct forms and suggests new ways to manipulate growth by targeting the molecular brakes rather than the hormone itself.
Citation: de Roij, M., Heijdra, E., Lamers, J. et al. Diversification of functional requirements for proteolysis of auxin response factors. Nat Commun 17, 3917 (2026). https://doi.org/10.1038/s41467-026-70440-0
Keywords: auxin signaling, protein degradation, Auxin Response Factors, Marchantia polymorpha, plant development