Clear Sky Science · de
Diversifizierung funktionaler Anforderungen an die Proteolyse von Auxin‑Antwortfaktoren
Wie Pflanzen ihr Wachstumshormon im Zaum halten
Pflanzen nutzen ein einfaches Molekül, das Hormon Auxin, um alles von Wurzeln und Blättern bis hin zu winzigen Fortpflanzungsstrukturen zu formen. Aus einem einfachen Signal die große Vielfalt pflanzlicher Formen entstehen zu lassen, erfordert jedoch enge Kontrolle. Diese Studie untersucht, wie Lebermoose ihre Empfindlichkeit gegenüber Auxin durch selektiven Abbau zentraler Regulationsproteine feinabstimmen und erklärt, warum einige dieser Proteine schnell abgebaut werden müssen, während andere gefahrlos länger vorhanden sein können.
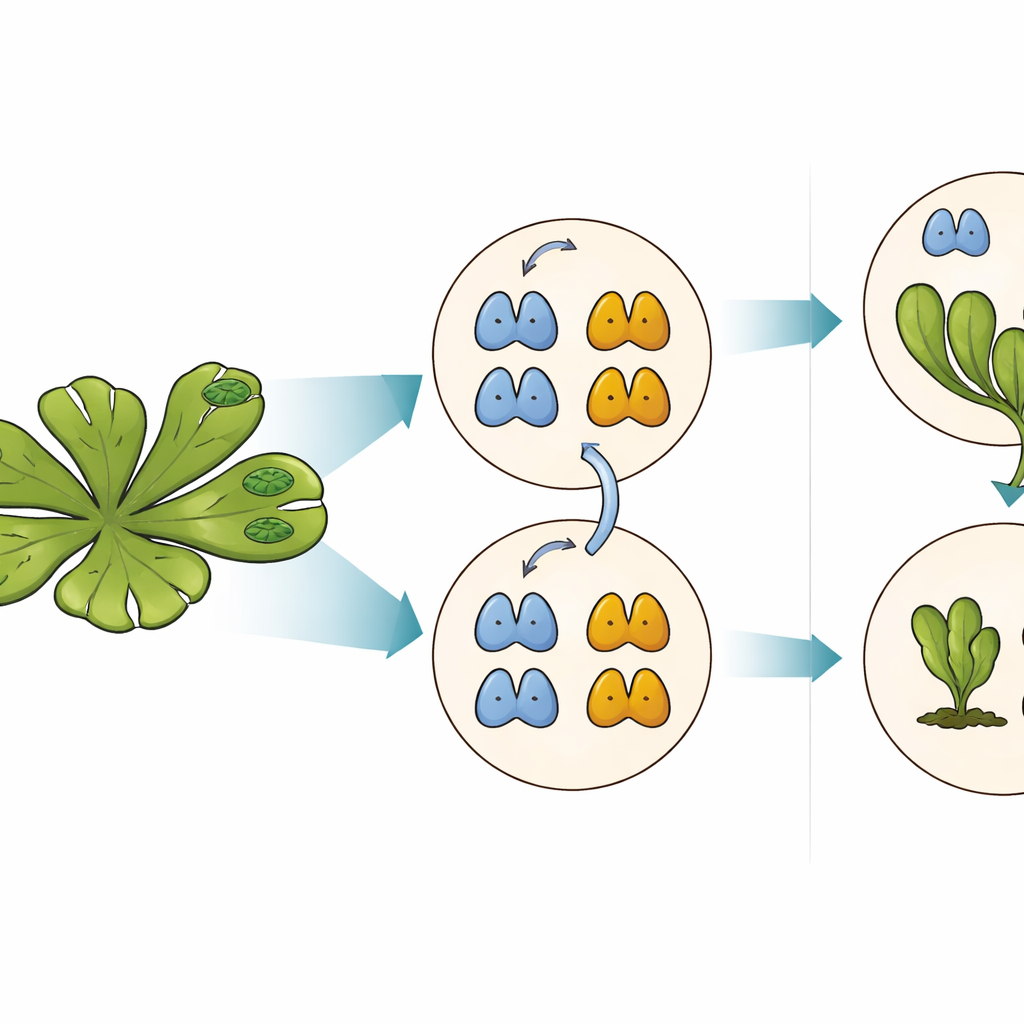
Das Gegengewicht hinter Auxin‑Reaktionen
Auxin wirkt größtenteils, indem es Gene über einen nukleären Signalweg in Pflanzenzellen an- oder ausschaltet. Im Zentrum dieses Weges stehen die Auxin‑Antwortfaktoren, oder ARFs, die an DNA binden und steuern, welche Gene auf das Hormon reagieren. Zwei Haupttypen von ARFs konkurrieren um dieselben DNA‑Bindungsstellen: A‑Klasse‑ARFs aktivieren in der Regel Gene als Reaktion auf Auxin, während B‑Klasse‑ARFs Gene abschalten und als Bremsen im System fungieren. Das Gleichgewicht zwischen diesen Aktivatoren und Repressoren in jeder Zelle bestimmt, wie stark die Zelle auf Auxin reagiert und damit, wie sich die Pflanze entwickelt.
Wenn Zerstörung zum Gestaltungselement wird
Jüngere Arbeiten zeigten, dass viele ARFs von Natur aus instabil sind: Sie tragen an ihrer DNA‑bindenden Spitze ein eingebautes Signal, das sie für den Abbau durch die zelluläre Proteinhydraulik, das Proteasom, markiert. Im Lebermoos Marchantia polymorpha ist jede ARF‑Klasse durch ein einzelnes Gen vertreten, wodurch sich dieses System sauber untersuchen lässt. Die Autoren veränderten systematisch einzelne Aminosäuren im Degradationsmotiv des B‑Klassen‑ARF MpARF2. Sie entdeckten, dass eine einzelne positiv geladene Baueinheit, eine Arginin‑Residue, entscheidend dafür ist, MpARF2 instabil zu machen. Die Entfernung seiner positiven Ladung oder das Umkehren benachbarter Ladungen stabilisiert das Protein stark, während viele andere Änderungen in derselben Region kaum Wirkung zeigen. Eine entsprechende Arginin‑Stelle im A‑Klassen‑Aktivator MpARF1 erfüllt eine ähnliche biochemische Rolle, was auf einen gemeinsamen, alten Ursprung dieses Degradationssignals hinweist.
Warum ein Protein schnell verschwinden muss und das andere bleiben kann
Obwohl sowohl MpARF1 als auch MpARF2 über dieses konservierte Motiv abgebaut werden können, unterscheidet sich ihr biologisches Abhängigkeitsverhältnis vom Abbau auffällig. Als die Forschenden MpARF1, den Aktivator, stabilisierten, stiegen die Proteinmengen nur mäßig, und Wachstum, Auxin‑Empfindlichkeit sowie Genexpressionsmuster blieben weitgehend normal. Im Gegensatz dazu hatte die Stabilisierung von MpARF2, dem Repressor, dramatische Folgen. Mithilfe fluoreszenter Markierungen und Proteasom‑Inhibitoren kartierten die Autoren den Abbau von MpARF2 über den Lebenszyklus der Pflanze. Sie fanden heraus, dass MpARF2 in normalen Pflanzen zwar in vielen Geweben gebildet wird, aber schnell entfernt wird, besonders in Wachstumszonen und rhizoidbildenden Zellen. Wenn der Abbau blockiert war, akkumulierte MpARF2 nahezu überall, und die Pflanzen zeigten schwere Defekte bei der Bildung flacher Thalli, Luftporen, Gemmabechern und Fortpflanzungsstrukturen.
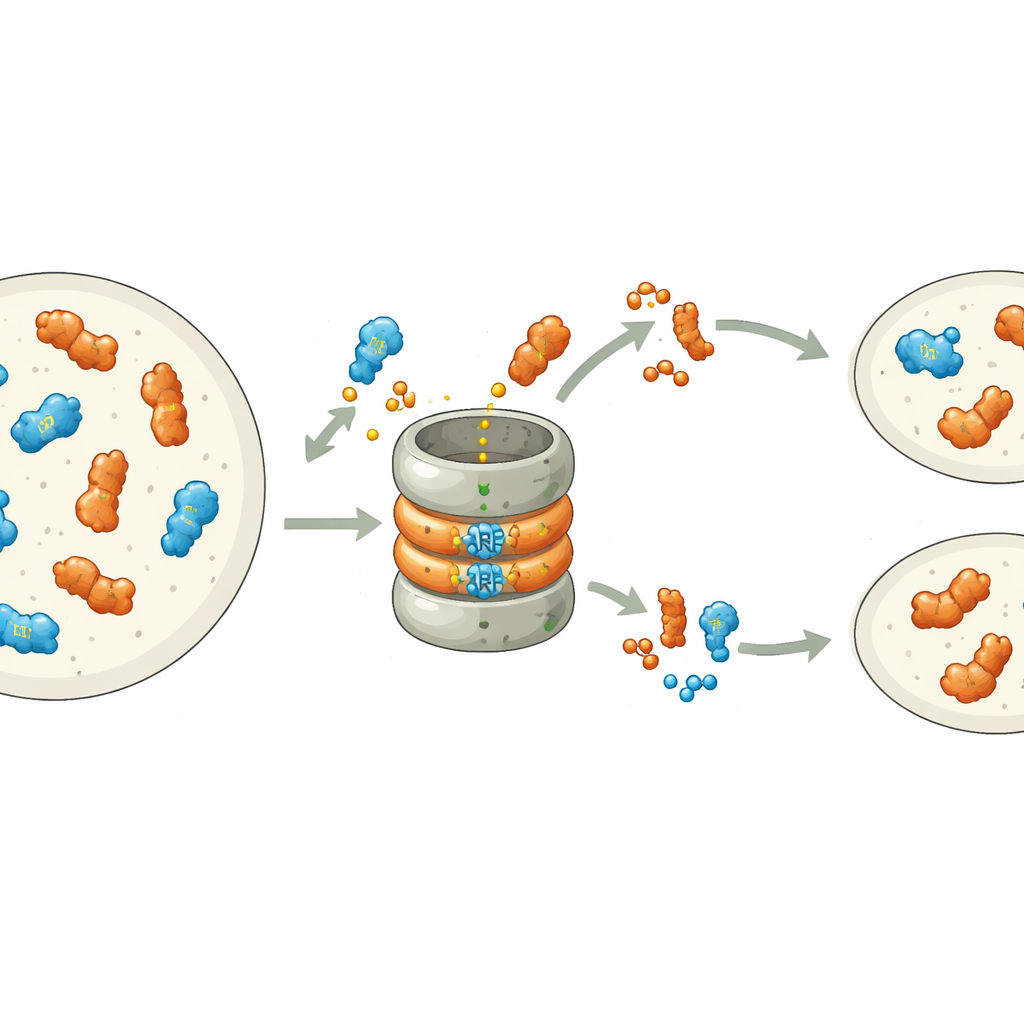
Lebenszyklus unter Kontrolle einer molekularen Bremse
Um Proteinstabilität direkt mit Funktion zu verknüpfen, kombinierten die Autoren genetische Mutanten mit clever konstruierten, schaltbaren Versionen von MpARF2. Das gezielte Einschalten einer degradationsresistenten MpARF2 reduzierte die Reaktion der Pflanzen auf zugegebenes Auxin deutlich und verhinderte die normale Entwicklung von Gemmabechern und männlichen Fortpflanzungsstielen. Die Ergänzung eines starken Verlust‑von‑Funktion‑Mutanten mit einem normalen MpARF2 stellte Wachstum und auxinabhängige Genaktivität teilweise wieder her, während eine stabilisierte Version dies nicht tat. Genomeweite RNA‑Sequenzierung bestätigte, dass typische auxin‑responsible Gene nicht reagieren, wenn MpARF2 nicht abgebaut werden kann, während viele nicht verwandte Gene fehlreguliert sind. Kurz gesagt: MpARF2 erfüllt seine richtige Rolle nur, wenn die Zelle seine Menge kontinuierlich durch Proteolyse trimmen kann.
Was das für Pflanzenvielfalt und Kontrolle bedeutet
Die Arbeit zeigt, dass trotz eines gemeinsamen alten Degradationssignals A‑ und B‑Klassen‑ARFs heute sehr unterschiedlich davon abhängig sind, zerstört zu werden. Der Aktivator MpARF1 verträgt reduzierten Abbau offenbar ohne große Folgen, möglicherweise weil andere Schutzmechanismen, wie separate Repressorproteine, seine Aktivität kontrollieren. Der Repressor MpARF2 hingegen muss kontinuierlich entfernt werden, um ein Überwältigen des Auxin‑Systems zu verhindern. Diese Divergenz hat sich wahrscheinlich während der Landbepflanzung entwickelt und erlaubt es einem einfachen Hormon, fein abgestimmte und gewebe‑spezifische Entwicklungsprogramme zu steuern. Indem die Studie zeigt, wie selektiver Proteinabbau Hormonantworten formt, hilft sie zu erklären, wie Pflanzen ein gemeinsames chemisches Signal in viele unterschiedliche Formen übersetzen, und schlägt neue Wege vor, Wachstum zu manipulieren, indem man die molekularen Bremsen statt des Hormons selbst anvisiert.
Zitation: de Roij, M., Heijdra, E., Lamers, J. et al. Diversification of functional requirements for proteolysis of auxin response factors. Nat Commun 17, 3917 (2026). https://doi.org/10.1038/s41467-026-70440-0
Schlüsselwörter: Auxin‑Signalgebung, Proteinabbau, Auxin‑Antwortfaktoren, Marchantia polymorpha, Pflanzenentwicklung