Clear Sky Science · it
Diversificazione dei requisiti funzionali per la proteolisi dei fattori di risposta all'auxina
Come le piante controllano il loro ormone di crescita
Le piante si affidano a una molecola semplice, l'ormone auxina, per plasmare tutto, dalle radici e dalle foglie fino alle piccole strutture riproduttive. Ma trasformare un segnale elementare nella ricca varietà di forme vegetali che osserviamo in natura richiede un controllo rigoroso. Questo studio esplora come le marchantie regolano finemente la loro sensibilità all'auxina distruggendo selettivamente proteine regolatorie chiave, rivelando perché alcune di queste proteine devono essere demolite rapidamente mentre altre possono rimanere più a lungo.
La squadra “spingi–tira” dietro le risposte all'auxina
L'auxina agisce principalmente attivando o spegnendo geni tramite una via nucleare all'interno delle cellule vegetali. Al centro di questa via ci sono i Fattori di Risposta all'Auxina, o ARF, che si legano al DNA e controllano quali geni rispondono all'ormone. Due tipi principali di ARF competono per gli stessi siti sul DNA: gli ARF di classe A in genere attivano i geni in risposta all'auxina, mentre gli ARF di classe B reprimono i geni e fungono da freni del sistema. L'equilibrio tra questi attivatori e repressori in ciascuna cellula determina quanto intensamente quella cellula reagisce all'auxina e, di conseguenza, come si sviluppa la pianta.
Quando la distruzione diventa un elemento progettuale
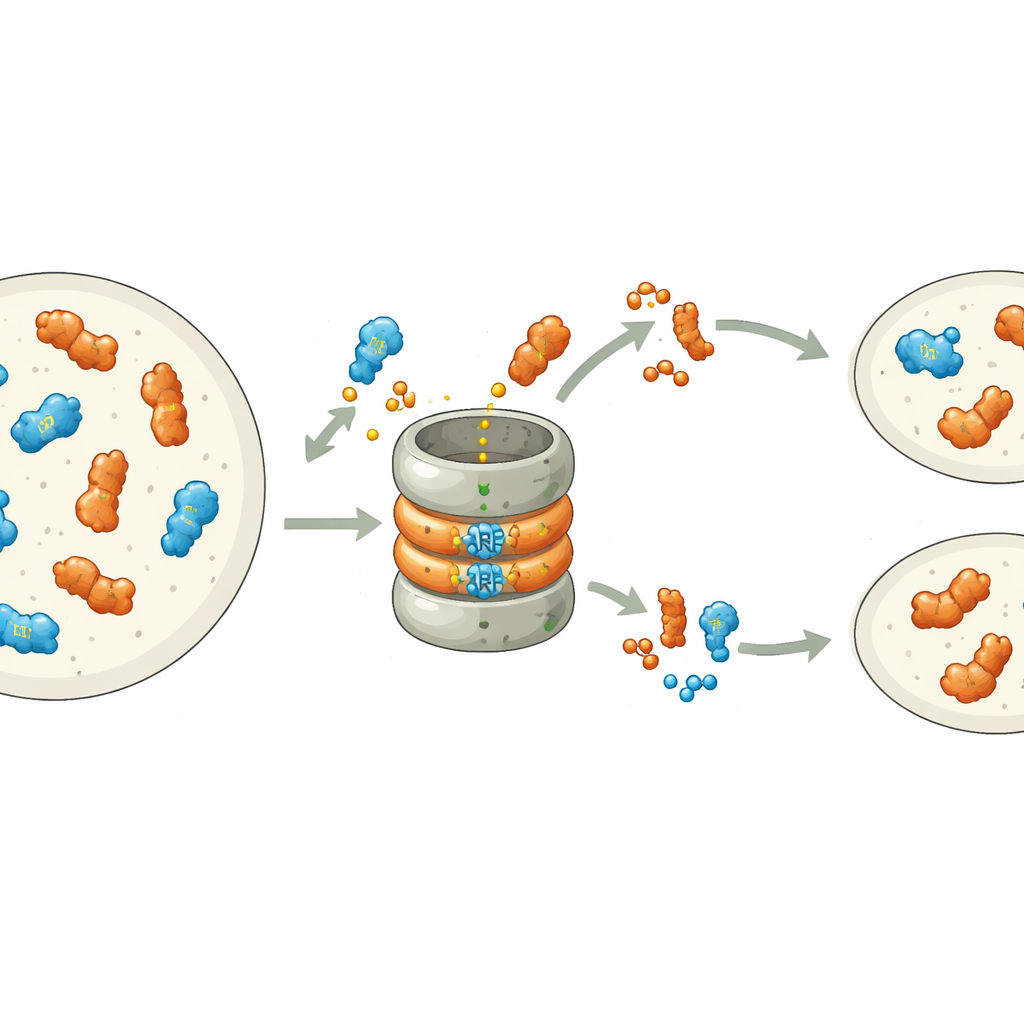
Lavori recenti hanno mostrato che molti ARF sono intrinsecamente instabili: portano un segnale incorporato nella loro estremità legante il DNA che li marca per la distruzione da parte della macchina cellulare che degrada le proteine, il proteasoma. In Marchantia polymorpha, ogni classe di ARF è rappresentata da un singolo gene, rendendo il sistema ideale per studiare questo processo. Gli autori hanno modificato sistematicamente singoli amminoacidi nel motivo di degradazione dell'ARF di classe B, MpARF2. Hanno scoperto che un singolo elemento carico positivamente, un arginina, è fondamentale per rendere MpARF2 instabile. Rimuovere la sua carica positiva o invertire le cariche vicine stabilizza fortemente la proteina, mentre molte altre modifiche nella stessa regione hanno scarso effetto. Un'arginina corrispondente nell'attivatore di classe A, MpARF1, svolge un ruolo biochimico simile, suggerendo un'origine antica condivisa di questo segnale di degradazione.
Perché una proteina deve scomparire rapidamente e l'altra può persistere
Sebbene sia MpARF1 sia MpARF2 possano essere degradati tramite questo motivo conservato, la loro dipendenza biologica dalla degradazione è sorprendentemente diversa. Quando i ricercatori hanno stabilizzato MpARF1, l'attivatore, i livelli proteici sono aumentati modestamente, ma la crescita della pianta, la sensibilità all'auxina e i modelli di espressione genica sono rimasti per lo più normali. Al contrario, stabilizzare MpARF2, il repressore, ha avuto conseguenze drastiche. Utilizzando marcatori fluorescenti e inibitori del proteasoma, il gruppo ha mappato la degradazione di MpARF2 lungo il ciclo di vita della pianta. Hanno trovato che nelle piante normali MpARF2 è prodotto in molti tessuti ma viene rapidamente rimosso, specialmente nelle punte di crescita e nelle cellule che formano i rizoidi. Quando la degradazione è stata bloccata, MpARF2 si è accumulato quasi ovunque e le piante hanno mostrato gravi difetti nella formazione di talli piatti, pori d'aria, coppe di gemme e strutture riproduttive.
Ciclo di vita sotto il controllo di un freno molecolare
Per collegare direttamente la stabilità proteica alla funzione, gli autori hanno combinato mutanti genetici con versioni ingegnerizzate e commutabili di MpARF2. Attivare una versione di MpARF2 resistente alla degradazione in momenti scelti riduceva nettamente la risposta delle piante all'auxina aggiunta e impediva lo sviluppo normale delle coppe di gemme e dei peduncoli riproduttivi maschili. Il complemento di un forte mutante loss‑of‑function con un MpARF2 normale ha ripristinato parzialmente la crescita e l'attività genica indotta dall'auxina, mentre una versione stabilizzata non lo ha fatto. L'analisi di RNA su scala genomica ha confermato che quando MpARF2 non può essere degradato, i geni tipicamente rispondenti all'auxina non reagiscono, mentre molti geni non correlati risultano malregolati. In sintesi, MpARF2 svolge il suo ruolo corretto solo quando la cellula può continuamente ridurne i livelli tramite proteolisi.
Cosa significa per la diversità e il controllo delle piante
Il lavoro rivela che, nonostante condividano un segnale di degradazione antico, gli ARF di classe A e B dipendono oggi in modo molto diverso dall'essere distrutti. L'attivatore MpARF1 può tollerare una ridotta degradazione con scarso impatto, probabilmente perché altre salvaguardie, come proteine repressorie separate, mantengono sotto controllo la sua attività. Il repressore MpARF2, invece, deve essere rimosso continuamente per evitare di sovraccaricare il sistema dell'auxina. Questa divergenza si è probabilmente evoluta quando le piante hanno colonizzato la terra, permettendo a un ormone semplice di guidare programmi di sviluppo finemente tarati e specifici per tessuto. Mostrando come la distruzione selettiva delle proteine scolpisca le risposte ormonali, lo studio aiuta a spiegare come le piante traducano un segnale chimico comune in molte forme distinte e suggerisce nuovi modi per manipolare la crescita mirando ai freni molecolari piuttosto che all'ormone stesso.
Citazione: de Roij, M., Heijdra, E., Lamers, J. et al. Diversification of functional requirements for proteolysis of auxin response factors. Nat Commun 17, 3917 (2026). https://doi.org/10.1038/s41467-026-70440-0
Parole chiave: segnalazione dell'auxina, degradazione proteica, Fattori di Risposta all'Auxina, Marchantia polymorpha, sviluppo delle piante