Clear Sky Science · pl
Różnicowanie wymagań funkcjonalnych dla proteolizy czynników odpowiedzi na auksynę
Jak rośliny kontrolują swój hormon wzrostu
Rośliny polegają na prostej cząsteczce, hormonie auksynie, by kształtować wszystko — od korzeni i liści po drobne struktury rozrodcze. Przekształcenie prostego sygnału w bogactwo form roślinnych, które widzimy w naturze, wymaga jednak ścisłej kontroli. Badanie to pokazuje, jak wątrobowce dopracowują swoją wrażliwość na auksynę przez selektywne niszczenie kluczowych białek regulacyjnych, wyjaśniając, dlaczego niektóre z nich muszą być szybko rozkładane, podczas gdy inne mogą bezpiecznie pozostawać w komórce.
Zespół popychu i hamowania stojący za reakcjami na auksynę
Auksyna działa głównie przez włączanie i wyłączanie genów za pośrednictwem jądrowej ścieżki wewnątrz komórek roślinnych. W centrum tej ścieżki znajdują się czynniki odpowiedzi na auksynę, czyli ARF-y, które wiążą DNA i kontrolują, które geny reagują na hormon. Dwa główne typy ARF-ów konkurują o te same miejsca na DNA: ARF-y klasy A zazwyczaj aktywują geny w odpowiedzi na auksynę, podczas gdy ARF-y klasy B wyciszają geny i działają jak hamulec systemu. Równowaga między tymi aktywatorami i represorami w każdej komórce determinuje, jak silnie ta komórka reaguje na auksynę, a w konsekwencji — jak rozwija się roślina.
Kiedy destrukcja staje się elementem projektu
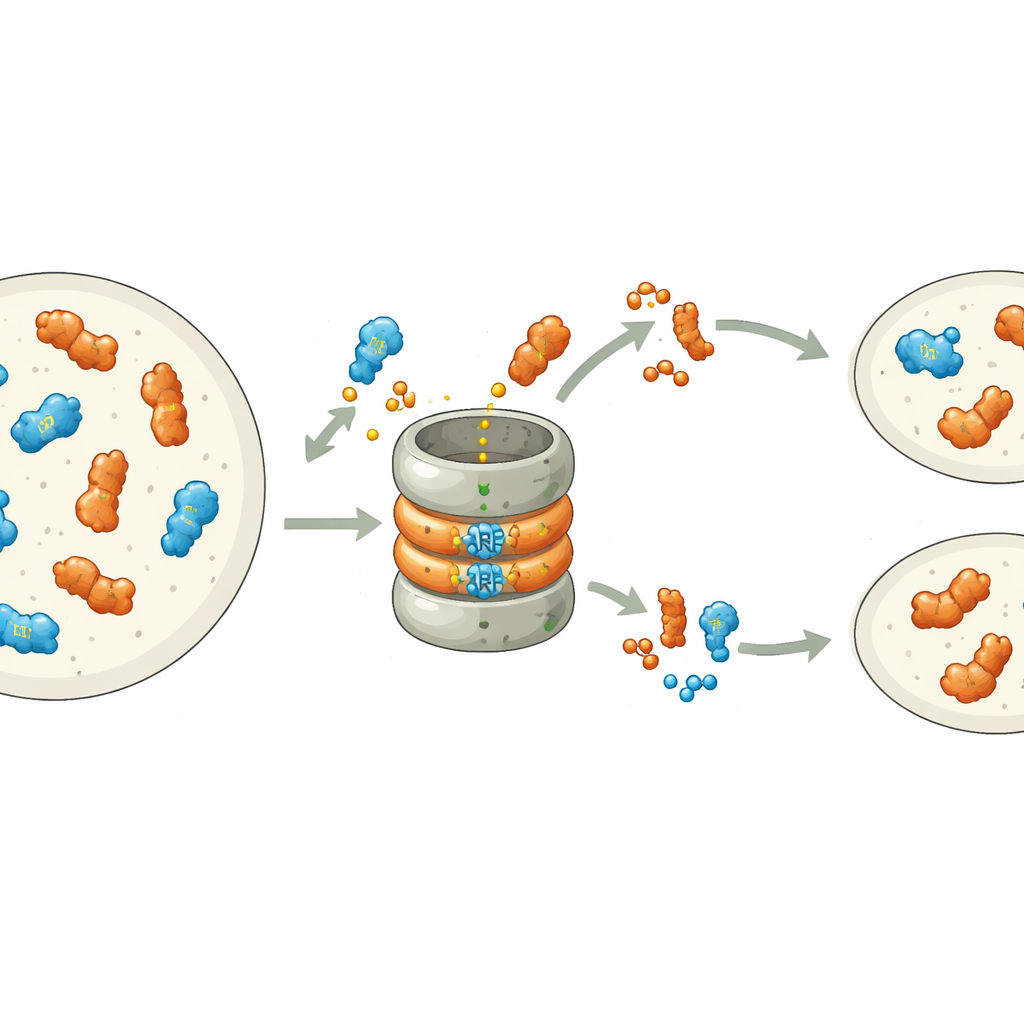
Wcześniejsze badania wykazały, że wiele ARF-ów jest z natury niestabilnych: niosą wbudowany sygnał w swojej końcówce wiążącej DNA, który oznacza je do degradacji przez komórkowe mechanizmy rozdrabniające białka, czyli proteasom. W wątrobowcu Marchantia polymorpha każda klasa ARF-ów reprezentowana jest przez pojedynczy gen, co tworzy przejrzysty system do badania tego procesu. Autorzy systematycznie zmieniali pojedyncze aminokwasy w motywie degradowalnym represora klasy B, MpARF2. Odkryli, że pojedynczy dodatnio naładowany element budulcowy — arginina — jest kluczowy dla niestabilności MpARF2. Usunięcie jej ładunku dodatniego lub odwrócenie sąsiednich ładunków silnie stabilizuje białko, podczas gdy wiele innych zmian w tym samym regionie ma niewielki efekt. Odpowiadająca arginina w aktywatorze klasy A, MpARF1, pełni podobną rolę biochemiczną, co wskazuje na wspólne, dawne pochodzenie tego sygnału degradacji.
Dlaczego jedno białko musi szybko zniknąć, a drugie może przetrwać
Chociaż zarówno MpARF1, jak i MpARF2 mogą być degradawane poprzez ten zachowany motyw, ich biologiczne uzależnienie od degradacji jest zasadniczo różne. Stabilizacja MpARF1, aktywatora, powodowała umiarkowany wzrost poziomów białka, lecz wzrost roślin, wrażliwość na auksynę i wzorce ekspresji genów pozostały w dużej mierze normalne. Natomiast stabilizacja MpARF2, represora, przyniosła dramatyczne konsekwencje. Dzięki zastosowaniu znaczników fluorescencyjnych i inhibitorów proteasomu zespół zmapował rozkład MpARF2 w całym cyklu życiowym rośliny. Stwierdzili, że w normalnych roślinach MpARF2 jest produkowany w wielu tkankach, ale szybko usuwany, szczególnie w stożkach wzrostu i komórkach tworzących ryzoidy. Gdy degradacja została zablokowana, MpARF2 kumulował się niemal wszędzie, a rośliny wykazywały poważne defekty w tworzeniu płaskich ciałek (thalli), porów powietrznych, kubków gemmowych i struktur rozrodczych.
Cykl życiowy pod kontrolą molekularnego hamulca
Aby powiązać stabilność białka bezpośrednio z funkcją, autorzy połączyli mutacje genetyczne z pomysłowo skonstruowanymi, przełączalnymi wersjami MpARF2. Włączenie odpornej na degradację wersji MpARF2 w wybranych momentach ostro zmniejszało odpowiedź roślin na dodaną auksynę i uniemożliwiało prawidłowy rozwój kubków gemmowych oraz męskich pędów rozrodczych. Uzupełnienie silnego mutanta o utracie funkcji normalnym MpARF2 częściowo przywracało wzrost i aktywność genów reagujących na auksynę, podczas gdy wersja ustabilizowana tego nie robiła. Sekwencjonowanie RNA obejmujące cały genom potwierdziło, że kiedy MpARF2 nie może być degradowany, typowe geny reagujące na auksynę nie reagują, natomiast wiele niepowiązanych genów wykazuje nieprawidłową regulację. Krótko mówiąc, MpARF2 pełni swoją właściwą rolę tylko wtedy, gdy komórka może ciągle przycinać jego poziomy poprzez proteolizę.
Co to oznacza dla różnorodności i kontroli roślin
Badanie ujawnia, że pomimo wspólnego, prastarego sygnału degradacji, ARF-y klasy A i B mają dziś bardzo różne zależności od bycia niszczonymi. Aktywator MpARF1 toleruje zmniejszoną degradację bez większych konsekwencji, być może dlatego, że inne zabezpieczenia, takie jak odrębne białka represorowe, utrzymują jego aktywność pod kontrolą. Represor MpARF2 musi jednak być nieustannie usuwany, by nie zdominować systemu auksynowego. Ta dywergencja prawdopodobnie ewoluowała wraz z kolonizacją lądu przez rośliny, pozwalając prostemu hormonowi napędzać precyzyjnie dostrojone i tkankowo specyficzne programy rozwojowe. Pokazując, jak selektywne niszczenie białek kształtuje odpowiedzi hormonalne, badanie pomaga wyjaśnić, jak rośliny przekładają wspólny sygnał chemiczny na wiele odmiennych form i sugeruje nowe sposoby manipulowania wzrostem przez celowanie w molekularne hamulce zamiast samego hormonu.
Cytowanie: de Roij, M., Heijdra, E., Lamers, J. et al. Diversification of functional requirements for proteolysis of auxin response factors. Nat Commun 17, 3917 (2026). https://doi.org/10.1038/s41467-026-70440-0
Słowa kluczowe: sygnalizacja auksynowa, degradacja białek, czynniki odpowiedzi na auksynę (ARF), Marchantia polymorpha, roślinny rozwój