Clear Sky Science · pt
Sinalização química de longo alcance in vivo é regulada por sinais mecânicos
Como o toque molda cérebros em crescimento
Enquanto um cérebro se forma, bilhões de neurônios jovens precisam navegar por caminhos longos e sinuosos para encontrar seus pares corretos. Há décadas, os cientistas sabem que essas jornadas são guiadas por sinais químicos que se espalham pelo tecido como odores invisíveis. Este estudo revela um parceiro surpreendente desses sinais químicos: a rigidez física do próprio cérebro. Ao sondar cuidadosamente cérebros de girinos em desenvolvimento, os autores mostram que a firmeza ou maciez do tecido pode ativar ou desativar sinais químicos distantes, direcionando finalmente as fibras nervosas em crescimento pelas rotas corretas.
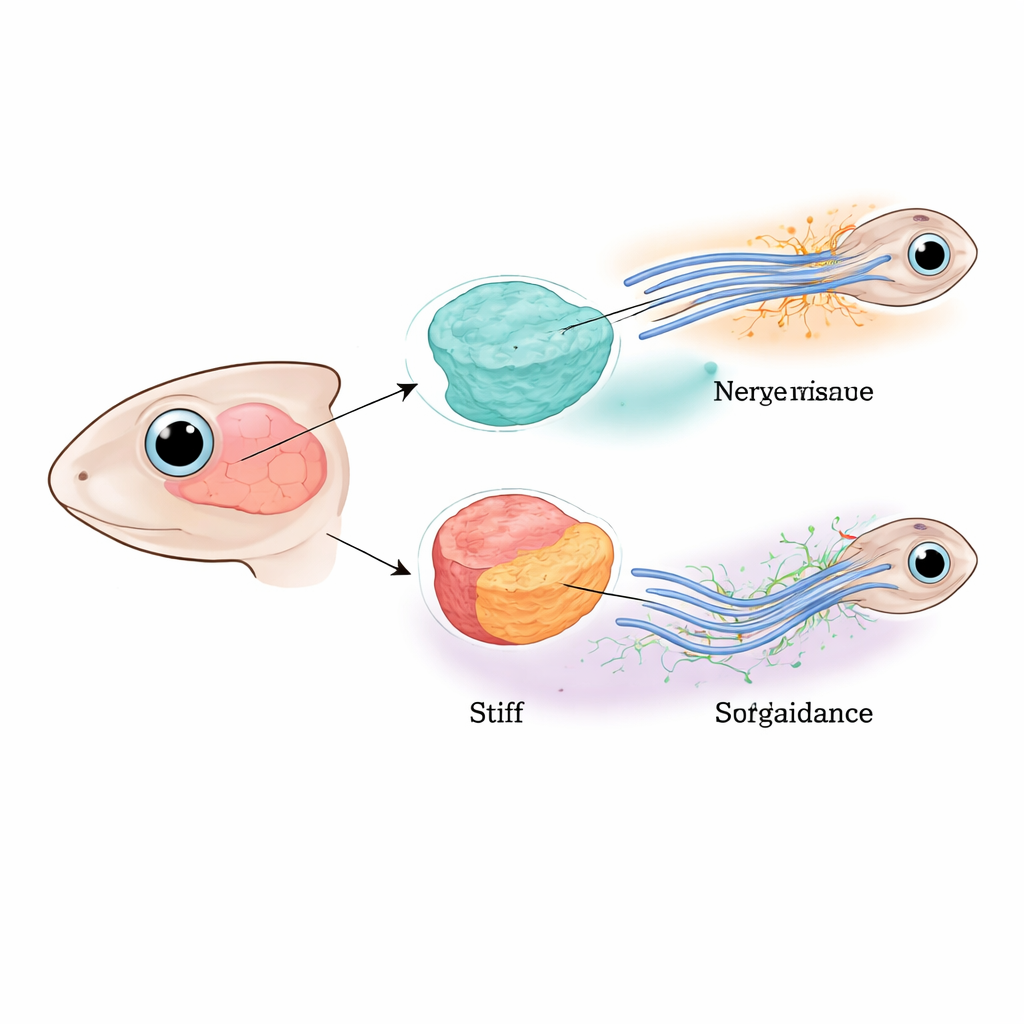
Guiando nervos através do cérebro jovem
Os pesquisadores concentraram-se na via óptica de girinos da rã africana Xenopus, onde as fibras nervosas do olho precisam atravessar o cérebro e executar uma curva precisa antes de alcançar sua região-alvo. Trabalhos anteriores mostraram que essas fibras são guiadas tanto por repelentes químicos difusíveis quanto por uma mudança gradual na rigidez do tecido ao longo de seu trajeto. Aqui, a equipe separou os papéis dos próprios neurônios e do tecido cerebral ao redor reduzindo seletivamente uma proteína-chave “sensora de força” chamada Piezo1 nas fibras nervosas, nas células cerebrais vizinhas ou em ambos. Quando Piezo1 estava ausente em todo lugar, muitas fibras estagnaram, se dividiram ou desviaram do percurso, confirmando que a percepção mecânica adequada é essencial para a fiação do cérebro.
Quando o cérebro amolece, sinais de longa distância enfraquecem
Em seguida, os autores investigaram como o tecido cerebral circundante contribui para esse sistema de orientação. Quando removeram Piezo1 apenas das fibras nervosas, a rigidez do tecido permaneceu normal, mas erros de navegação ainda ocorreram, indicando que as fibras não conseguiam mais sentir adequadamente seu ambiente mecânico. De forma marcante, quando Piezo1 foi removido do tecido cerebral, o tecido tornou-se quase duas vezes mais mole, e os erros de orientação tornaram-se ainda mais frequentes — mesmo que as fibras nervosas ainda contivessem Piezo1. Nessas regiões amolecidas, os níveis de duas moléculas conhecidas de orientação de longo alcance, Slit1 e Semaphorin3A, caíram acentuadamente tanto no RNA quanto no nível proteico. Isso mostrou que o estado físico do cérebro não era apenas sentido passivamente pelas células; ele controlava ativamente quanto desses sinais químicos difusíveis eram produzidos.
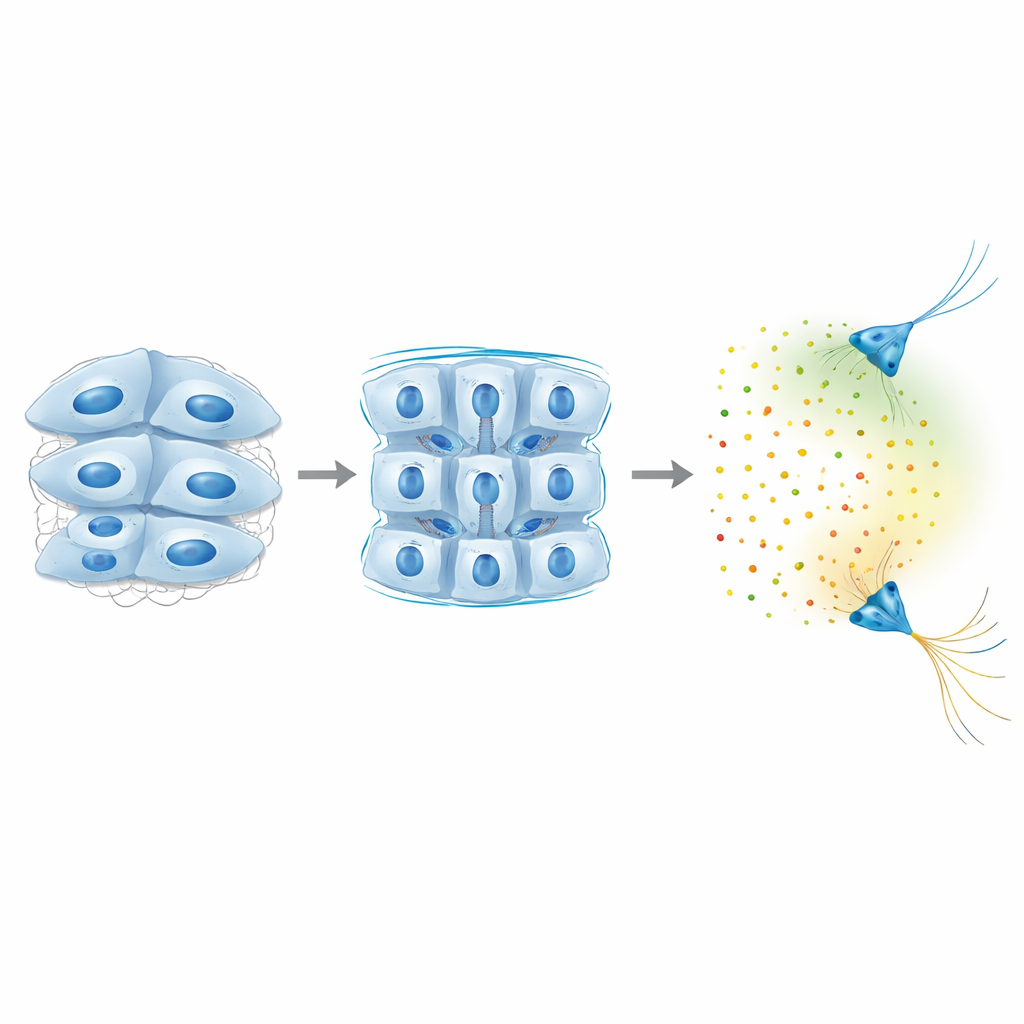
A adesão celular como uma alavanca mecânica oculta
O que torna o tecido cerebral deficiente em Piezo1 mais macio? A equipe descartou explicações simples, como menos células ou células individuais mais flácidas. Em vez disso, descobriram que proteínas importantes que colam células cerebrais vizinhas — NCAM1 e N‑cadherina — estavam reduzidas quando os níveis de Piezo1 caíam. Quando os pesquisadores diminuíram diretamente essas proteínas de adesão, a rigidez tecidual diminuiu e os níveis de Semaphorin3A também caíram, mesmo que o interior das células permanecesse mecanicamente inalterado. Isso aponta para uma cadeia de eventos em que a atividade de Piezo1 ajuda a manter fortes conexões célula–célula, que por sua vez definem a rigidez global do tecido, a qual então governa quanto de certos sinais químicos de longo alcance o tecido produz.
Girando o botão mecânico para cima
Para testar o outro lado dessa relação, os autores aumentaram a rigidez cerebral de várias maneiras. Em placas de cultura, eles embutiram pequenos pedaços de tecido cerebral macio em géis 3D macios ou rígidos. Em géis mais rígidos, o tecido puxou com mais força seu entorno e passou a produzir níveis muito maiores de Slit1 e Semaphorin3A, mesmo em regiões que normalmente não produzem esses sinais. Em girinos intactos, eles aplicaram um fármaco que aumenta as forças contráteis internas ou comprimiram suavemente regiões cerebrais específicas com uma sonda minúscula. Ambas as abordagens aumentaram a rigidez nas regiões escolhidas e desencadearam nova produção de Semaphorin3A em áreas que costumam ser quimicamente silenciosas. Contudo, esse “interruptor” mecânico falhou em animais nos quais Piezo1 havia sido reduzido, mostrando que o canal é necessário para traduzir rigidez adicional em sinalização química aumentada.
Por que isso importa para cérebros em crescimento e doentes
No conjunto, o estudo revela um circuito de retroalimentação no qual as células cerebrais puxam e sentem seu entorno por meio de Piezo1, ajustam suas conexões adesivas para definir a rigidez do tecido e, somente uma vez que um limiar de rigidez é alcançado, ativam sinais químicos de longo alcance que orientam fibras nervosas distantes. Em vez de agirem independentemente, sinais mecânicos e químicos estão intimamente entrelaçados: uma mudança local na firmeza do tecido pode remodelar o “clima químico” em lugares distantes, influenciando onde os axônios crescem e como os circuitos se formam. Essa percepção sugere que a rigidez tecidual alterada — observada em condições que vão de lesão cerebral à neurodegeneração — pode repercutir por cérebros em desenvolvimento ou envelhecidos ao alterar os próprios sinais que as células usam para se comunicar a longa distância.
Citação: Pillai, E.K., Mukherjee, S., Gampl, N. et al. Long-range chemical signalling in vivo is regulated by mechanical signals. Nat. Mater. 25, 687–697 (2026). https://doi.org/10.1038/s41563-025-02463-9
Palavras-chave: mecanotransdução, guia de axônio, rigidez tecidual, Piezo1, desenvolvimento cerebral