Clear Sky Science · zh
半胱氨酸供应通过LRRC58 E3连接酶与其底物CDO1的相反稳定性调节泛素信号
细胞如何决定何时保留或丢弃一项酶
在每个细胞内部,蛋白分子不断被合成与降解。这种无声的周转帮助细胞适应变化的营养与压力。该研究揭示了人类细胞如何感知含硫氨基酸半胱氨酸的可用性,并通过精细配线的蛋白质处置系统调节一项关键代谢酶的水平。
一种指示“够了”的营养信号
半胱氨酸是细胞必须保持平衡的氨基酸。太少会威胁生命过程;太多也可能有害。细胞管理半胱氨酸的方式之一是通过一种称为半胱氨酸二氧化酶1(CDO1)的酶来分解它。早期动物研究显示,当半胱氨酸匮乏时,CDO1会被破坏;当半胱氨酸充足时,CDO1则被保留。然而,在低半胱氨酸条件下挑选出CDO1以便删除的分子机械尚未被明确定义。
寻找像营养调节器一样的蛋白
研究人员使用大规模蛋白筛选方法,比较有无半胱氨酸条件下的人类细胞。他们关注一类称为Cullin‑RING连接酶的蛋白复合体,这些复合体像可定制的钩子一样为选定蛋白加上被降解的标签。在半胱氨酸匮乏的细胞中,一个钩子成分——名为LRRC58的受体蛋白——格外突出:它在半胱氨酸缺乏时变得更丰富,并加入到活性的连接酶复合体中。与此同时,CDO1的水平骤降,提示LRRC58与CDO1之间存在相反的关联。在多种细胞类型中,当LRRC58在低半胱氨酸下水平升高时,CDO1几乎检测不到;当补充半胱氨酸时,LRRC58下降而CDO1恢复。
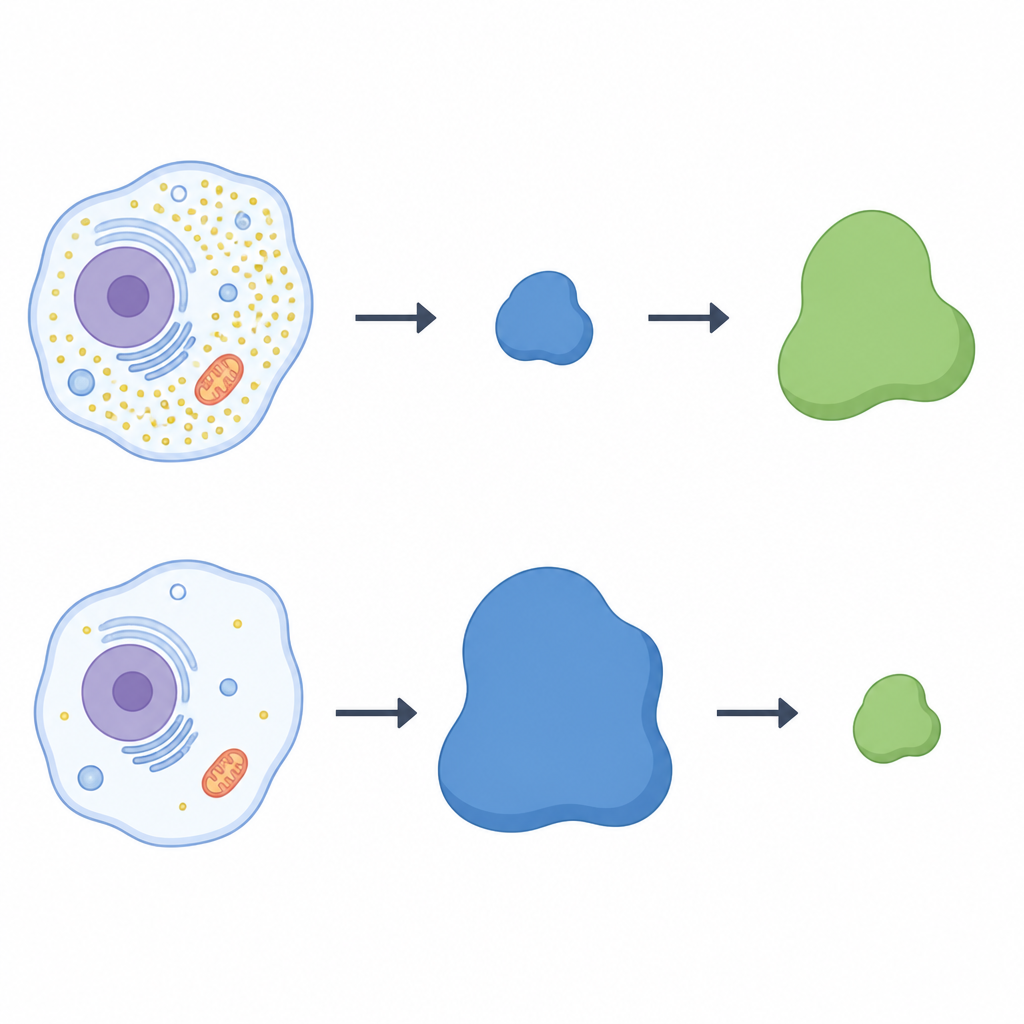
两种蛋白之间的天枰效应
为检验LRRC58是否直接控制CDO1,团队在人体细胞中敲除了LRRC58基因。没有LRRC58时,在半胱氨酸饥饿期间CDO1不再消失;即便半胱氨酸低,酶也持续存在。补回一个功能性LRRC58能恢复CDO1的丧失,而一个无法与其连接酶伙伴结合的LRRC58突变体则不能。荧光报告系统证实CDO1的稳定性依赖于LRRC58及特定的cullin‑RING骨架。综合这些实验证据表明,LRRC58与CDO1形成一对天枰:LRRC58通常不稳定,但在半胱氨酸耗尽时被稳定化,反过来在同样条件下将CDO1标记为待降解对象。
以原子级细节观察处置机器
作者从纯化蛋白重建了LRRC58系统,观察到LRRC58与基于cullin的骨架(CUL2或CUL5)配对,形成一个活性标记机器,将泛素分子附加到CDO1上。借助冷冻电子显微镜,他们获得了该机器运作的接近原子分辨率的图像。这些结构展示了LRRC58如何在若干接触面上抓住CDO1,并将CDO1上的单个氨基酸——赖氨酸8(K8)——定位到接近泛素转移化学位点的位置。突变赖氨酸8或削弱接触面会阻止CDO1在半胱氨酸短缺时的降解,证实结构中看到的几何构型对于系统在细胞内的功能至关重要。
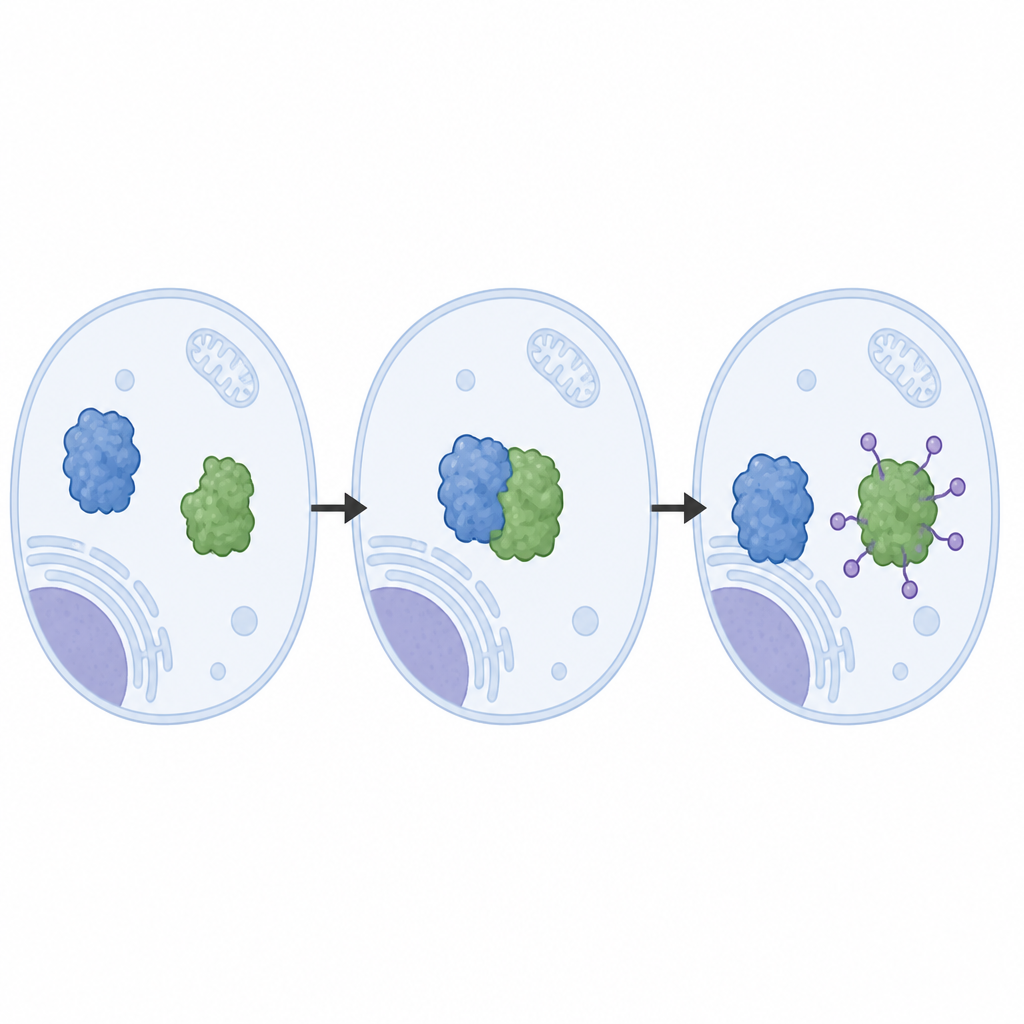
天然控制路径与药物诱导捷径的对比
研究还比较了这一天然通路与一种基于药物的策略。最近开发的一种“分子胶”化合物迫使CDO1粘连到另一受体蛋白VHL上,VHL使用相同类型的cullin骨架但以不同的取向抓持CDO1。在细胞与试管反应中,这种药物引导的途径在CDO1的多个位点打标签,而不仅仅是赖氨酸8,并且即便在疾病相关突变破坏LRRC58对其的正常识别时也能有效破坏CDO1。这一对比表明,药物可以利用细胞自身的处置硬件,同时绕开对标记位置与方式的一些天然限制。
这对细胞健康与未来疗法的意义
对非专业读者而言,核心信息是细胞使用精细调节的蛋白处置系统将酶水平与营养供应相匹配。当半胱氨酸匮乏时,LRRC58被稳定并与其伙伴组装,选择性地标记CDO1以便清除,帮助细胞节省半胱氨酸;当半胱氨酸充足时,LRRC58本身被降解,从而保护CDO1,使多余的半胱氨酸得以安全分解。通过描绘这一路径并以高分辨率捕获其结构,这项工作解释了代谢学中长期观察到的现象,并说明有针对性的药物未来如何通过将特定蛋白引导进入或绕过此类天然控制回路来救援或清除它们。
引用: Andree, G.A., Stier, L.J., Schmiederer, K. et al. Cysteine availability tunes ubiquitin signaling via inverse stability of LRRC58 E3 ligase and its substrate CDO1. Nat Commun 17, 4196 (2026). https://doi.org/10.1038/s41467-026-72524-3
关键词: 半胱氨酸代谢, 蛋白降解, 泛素系统, CDO1 酶, E3 连接酶