Clear Sky Science · en
Cysteine availability tunes ubiquitin signaling via inverse stability of LRRC58 E3 ligase and its substrate CDO1
How Cells Decide When to Save or Scrap an Enzyme
Inside every cell, protein molecules are constantly being built up and broken down. This silent churn helps cells adapt to changing diets and stresses. This study uncovers how human cells sense the availability of the sulfur‑containing amino acid cysteine and then fine‑tune the levels of a key metabolic enzyme, using a carefully wired protein disposal system.
A Nutrient That Signals When Enough Is Enough
Cysteine is an amino acid that cells must keep in balance. Too little threatens vital processes; too much can be harmful. One way cells manage cysteine is by breaking it down with an enzyme called cysteine dioxygenase 1, or CDO1. Earlier animal work showed that when cysteine is scarce, CDO1 is destroyed, and when cysteine is plentiful, CDO1 is preserved. However, the molecular machinery that picks out CDO1 for removal in low cysteine conditions had not been clearly identified.
Finding the Protein that Acts Like a Nutrient Dial
The researchers used a large‑scale protein screening method to scan human cells grown with or without cysteine. They focused on a family of protein complexes called cullin‑RING ligases, which act like customizable hooks that tag selected proteins for destruction. In cysteine‑starved cells, one hook component, a receptor protein named LRRC58, stood out: it became more abundant and joined active ligase complexes only when cysteine was lacking. At the same time, levels of CDO1 dropped sharply, suggesting an inverse link between LRRC58 and CDO1. Across several cell types, when LRRC58 levels were high under low cysteine, CDO1 was nearly undetectable; when cysteine was added back, LRRC58 fell and CDO1 rebounded.
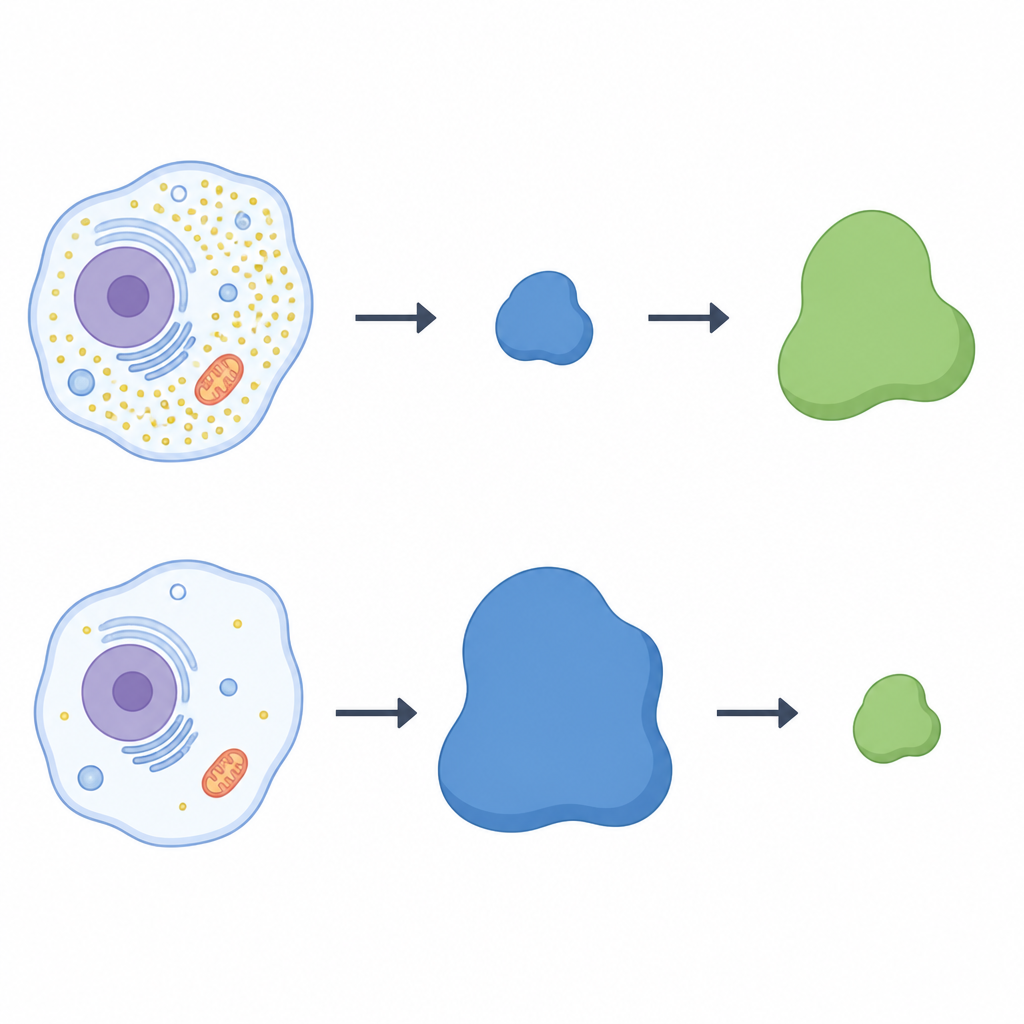
A See‑Saw Between Two Proteins
To test whether LRRC58 directly controls CDO1, the team disabled the LRRC58 gene in human cells. Without LRRC58, CDO1 no longer disappeared during cysteine starvation; the enzyme persisted even when cysteine was low. Adding back a working version of LRRC58 restored CDO1 loss, while a mutant form of LRRC58 that cannot join its ligase partners failed to do so. A fluorescent reporter system confirmed that CDO1’s stability depends on both LRRC58 and specific cullin‑RING scaffolds. Taken together, these experiments show that LRRC58 and CDO1 form a see‑saw pair: LRRC58 is normally unstable but becomes stabilized when cysteine runs low, and in turn it targets CDO1 for destruction under those same conditions.
Watching the Disposal Machine at Atomic Detail
The authors reconstructed the LRRC58 system from purified proteins and observed that LRRC58 pairs with cullin‑based scaffolds (CUL2 or CUL5) to form an active tagging machine that attaches ubiquitin molecules to CDO1. Using cryo‑electron microscopy, they obtained near‑atomic images of this machine in action. These structures reveal how LRRC58 grips CDO1 at several contact patches and positions a single amino acid on CDO1, lysine‑8, right next to the chemical site where ubiquitin is transferred. Mutating lysine‑8, or weakening the contact patches, blocks CDO1 break‑down in response to cysteine shortage, confirming that the geometry seen in the structures is essential for how the system works inside cells.
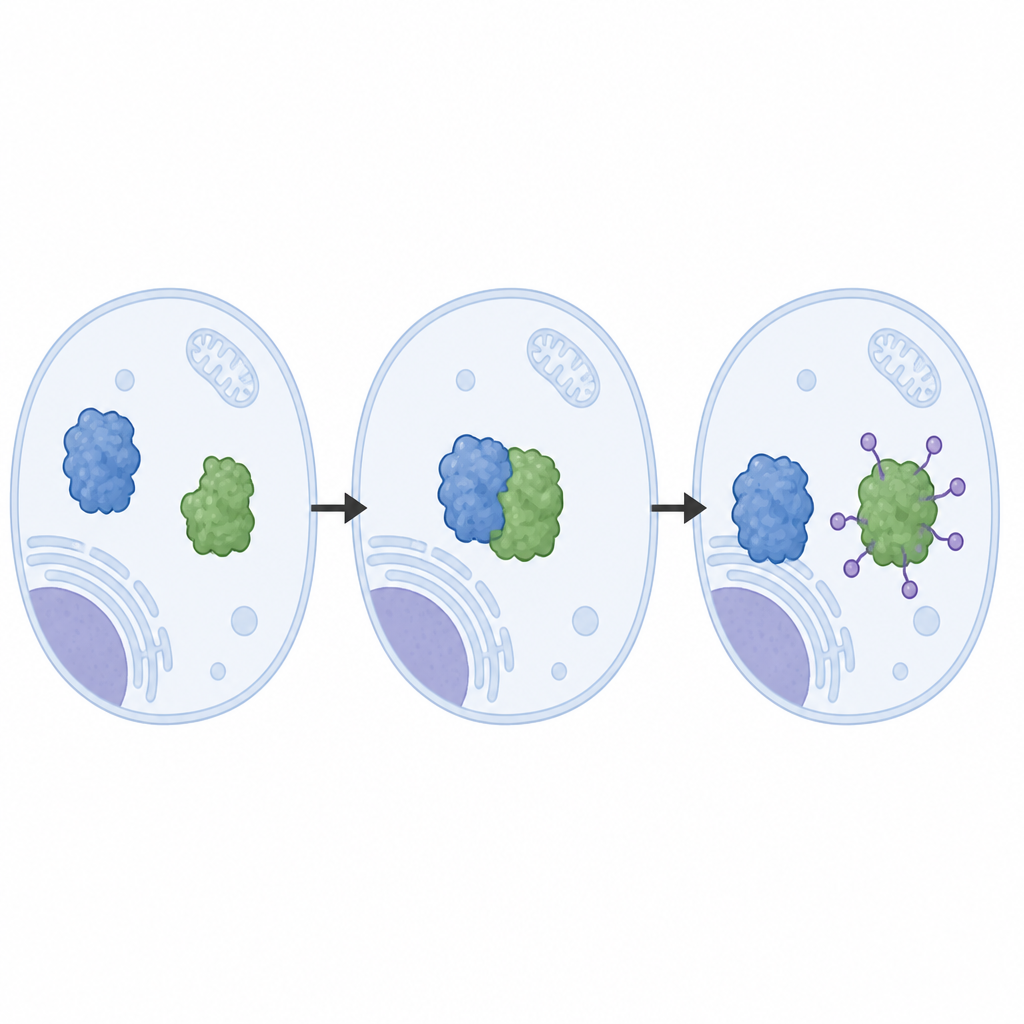
Natural Control Versus Drug‑Induced Shortcuts
The study also compares this natural pathway with a drug‑based strategy. A recently developed “molecular glue” compound forces CDO1 to stick to a different receptor protein called VHL, which uses the same type of cullin scaffold but holds CDO1 in another orientation. In cells and test‑tube reactions, this drug‑guided route tags many sites on CDO1, not just lysine‑8, and can efficiently destroy CDO1 even when disease‑linked mutations disrupt its normal recognition by LRRC58. This contrast shows how drugs can tap into the cell’s own disposal hardware while bypassing some of the natural constraints on where and how a protein is tagged.
What This Means for Cell Health and Future Therapies
For a non‑specialist, the key message is that cells use a finely tuned protein disposal system to match enzyme levels to nutrient supply. When cysteine is scarce, LRRC58 becomes stable, assembles with its partners, and selectively marks CDO1 for removal, helping the cell conserve cysteine. When cysteine is abundant, LRRC58 is itself degraded, sparing CDO1 so the excess cysteine can be safely broken down. By mapping this pathway and capturing it at high resolution, the work explains a long‑standing observation in metabolism and illustrates how targeted drugs might one day rescue or remove specific proteins by redirecting them into or around such natural control circuits.
Citation: Andree, G.A., Stier, L.J., Schmiederer, K. et al. Cysteine availability tunes ubiquitin signaling via inverse stability of LRRC58 E3 ligase and its substrate CDO1. Nat Commun 17, 4196 (2026). https://doi.org/10.1038/s41467-026-72524-3
Keywords: cysteine metabolism, protein degradation, ubiquitin system, CDO1 enzyme, E3 ligase