Clear Sky Science · fr
La disponibilité de la cystéine module la signalisation par ubiquitine via la stabilité inverse de la ligase E3 LRRC58 et de son substrat CDO1
Comment les cellules décident de conserver ou de jeter une enzyme
À l’intérieur de chaque cellule, les protéines sont en permanence synthétisées et dégradées. Ce va-et-vient discret aide les cellules à s’adapter à des régimes alimentaires et à des stress variables. Cette étude révèle comment les cellules humaines détectent la disponibilité de l’acide aminé soufré cystéine puis ajustent finement les niveaux d’une enzyme métabolique clé, en s’appuyant sur un système d’élimination protéique précisément câblé.
Un nutriment qui signale quand c’en est assez
La cystéine est un acide aminé dont la concentration doit être maintenue en équilibre. Trop peu menace des processus vitaux ; trop peut être toxique. Une façon pour la cellule de gérer la cystéine est de la dégrader grâce à une enzyme nommée cystéine dioxygénase 1, ou CDO1. Des travaux antérieurs sur des animaux montraient que lorsque la cystéine manque, CDO1 est détruite, et que lorsque la cystéine est abondante, CDO1 est préservée. Cependant, la machinerie moléculaire qui cible CDO1 pour élimination en condition de pénurie n’avait pas été clairement identifiée.
Identifier la protéine qui agit comme un réglage nutritionnel
Les chercheurs ont utilisé une méthode de criblage protéique à grande échelle pour analyser des cellules humaines cultivées avec ou sans cystéine. Ils se sont concentrés sur une famille de complexes protéiques appelés ligases cullin‑RING, qui fonctionnent comme des crochets modulables marquant des protéines sélectionnées pour destruction. Dans les cellules privées de cystéine, un composant de ces crochets, une protéine réceptrice appelée LRRC58, est apparu comme remarquable : elle devenait plus abondante et s’incorporait aux complexes de ligase actifs seulement en l’absence de cystéine. En parallèle, les niveaux de CDO1 chutaient fortement, suggérant un lien inverse entre LRRC58 et CDO1. Dans plusieurs types cellulaires, lorsque LRRC58 était élevé en situation de carence en cystéine, CDO1 était presque indétectable ; lorsque la cystéine était rétablie, LRRC58 diminuait et CDO1 revenait.
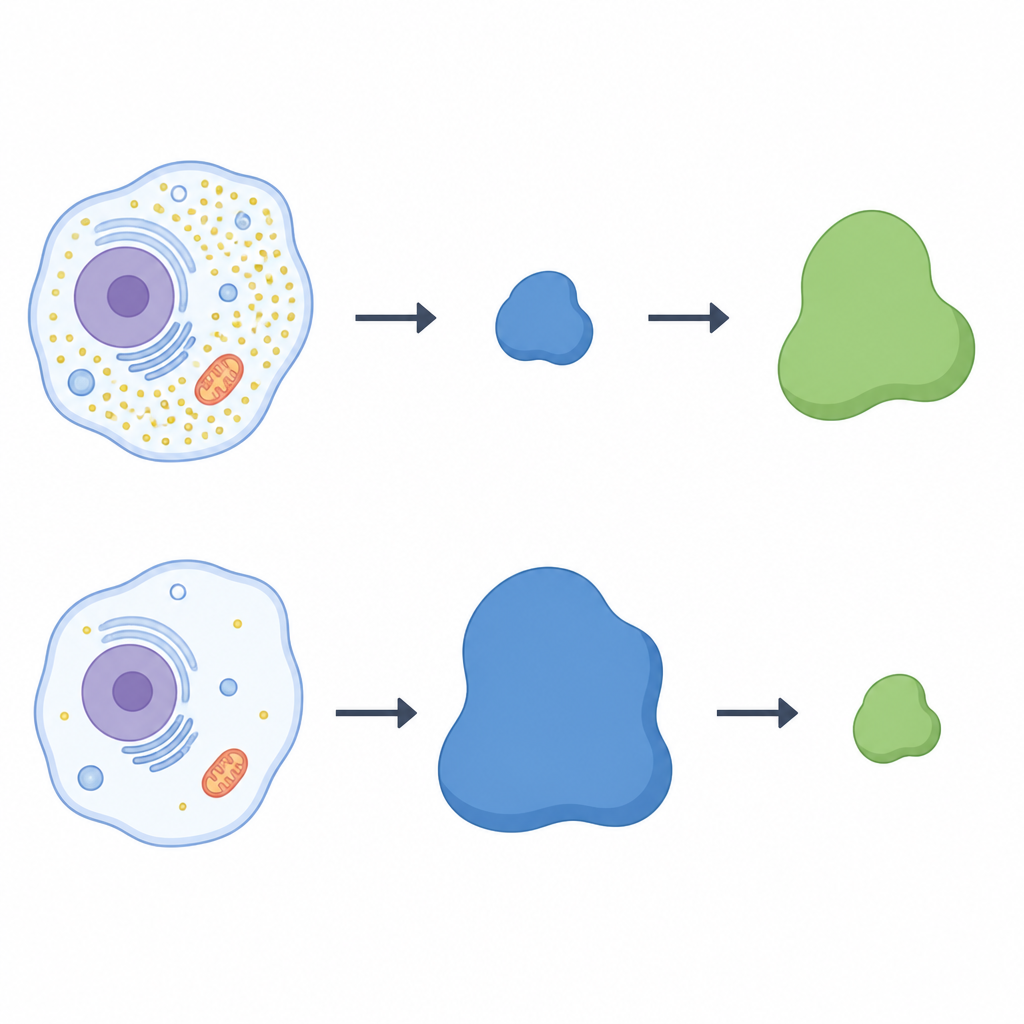
Une bascule entre deux protéines
Pour vérifier si LRRC58 contrôle directement CDO1, l’équipe a inactivé le gène LRRC58 dans des cellules humaines. Sans LRRC58, CDO1 ne disparaissait plus lors de la privation en cystéine ; l’enzyme persistait même lorsque la cystéine était faible. La réintroduction d’une version fonctionnelle de LRRC58 rétablit la perte de CDO1, tandis qu’une forme mutante de LRRC58 incapable de s’associer à ses partenaires de ligase ne réussissait pas à le faire. Un système rapporteur fluorescent a confirmé que la stabilité de CDO1 dépend à la fois de LRRC58 et de scaffolds cullin‑RING spécifiques. Pris ensemble, ces expériences montrent que LRRC58 et CDO1 forment une paire en équilibre : LRRC58 est normalement instable mais se stabilise lorsque la cystéine vient à manquer, et à son tour il cible CDO1 pour destruction dans ces mêmes conditions.
Observer la machine d’élimination au détail atomique
Les auteurs ont reconstruit le système LRRC58 à partir de protéines purifiées et ont observé que LRRC58 s’associe à des échafaudages basés sur cullin (CUL2 ou CUL5) pour former une machine active qui attache des molécules d’ubiquitine à CDO1. Grâce à la cryo‑microscopie électronique, ils ont obtenu des images quasi atomiques de cette machine en action. Ces structures montrent comment LRRC58 saisit CDO1 sur plusieurs points de contact et positionne un seul acide aminé de CDO1, la lysine‑8, juste à côté du site chimique où l’ubiquitine est transférée. La mutation de la lysine‑8, ou l’affaiblissement des points de contact, empêche la dégradation de CDO1 en réponse à la pénurie de cystéine, confirmant que la géométrie observée dans les structures est essentielle au fonctionnement du système dans les cellules.
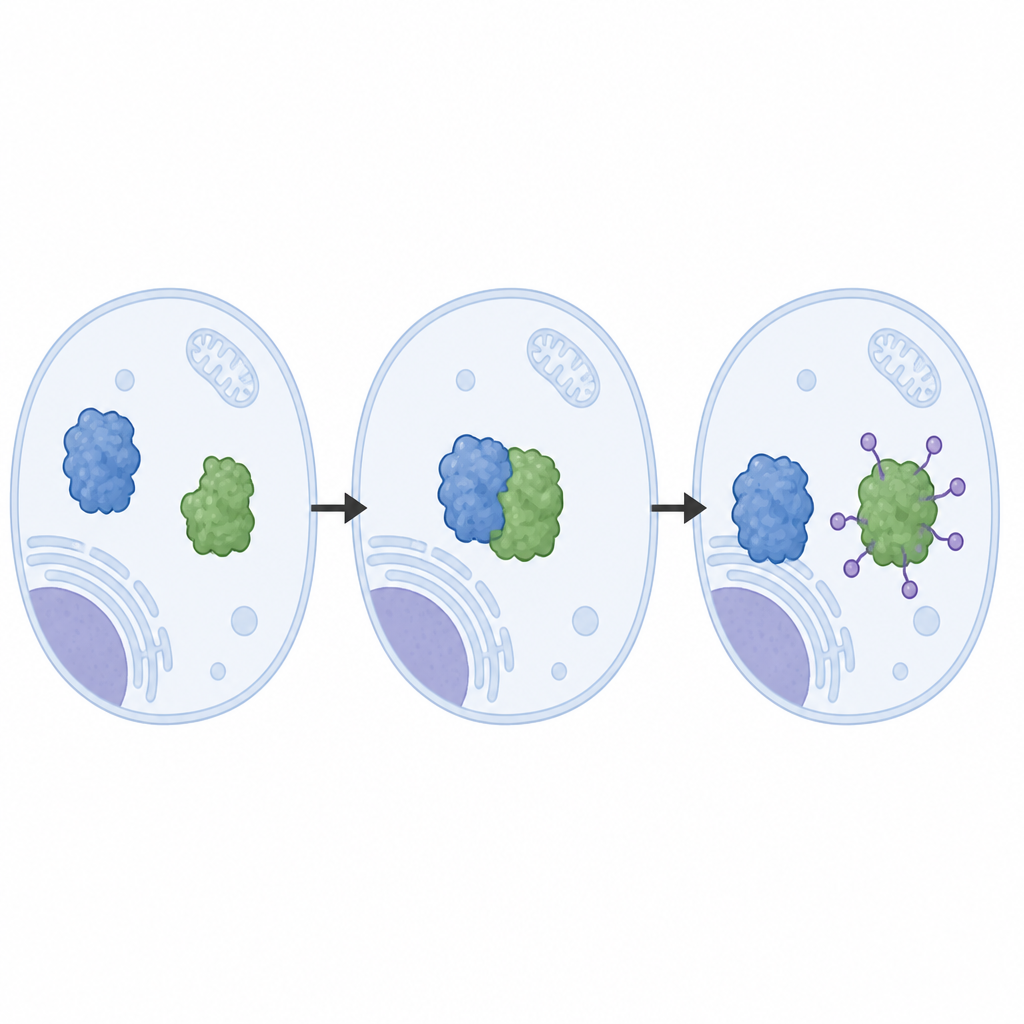
Contrôle naturel versus raccourcis induits par des médicaments
L’étude compare aussi cette voie naturelle à une stratégie fondée sur un médicament. Un composé récemment développé, appelé « colle moléculaire », force CDO1 à se lier à un autre récepteur nommé VHL, qui utilise le même type d’échafaudage cullin mais maintient CDO1 dans une orientation différente. Dans les cellules et les réactions in vitro, cette voie guidée par le médicament marque de nombreux sites sur CDO1, pas seulement la lysine‑8, et peut détruire efficacement CDO1 même lorsque des mutations liées à la maladie perturbent sa reconnaissance normale par LRRC58. Ce contraste montre comment des médicaments peuvent exploiter l’appareillage d’élimination de la cellule tout en contournant certaines contraintes naturelles sur l’emplacement et la manière dont une protéine est marquée.
Ce que cela signifie pour la santé cellulaire et les thérapies futures
Pour un public non spécialiste, le message clé est que les cellules utilisent un système d’élimination des protéines finement réglé pour adapter les niveaux enzymatiques à l’apport en nutriment. Lorsque la cystéine manque, LRRC58 se stabilise, s’assemble avec ses partenaires et marque sélectivement CDO1 pour élimination, aidant la cellule à conserver la cystéine. Lorsque la cystéine est abondante, LRRC58 est lui‑même dégradé, épargnant CDO1 afin que l’excès de cystéine puisse être dégradé en toute sécurité. En cartographiant cette voie et en la capturant à haute résolution, ce travail explique une observation ancienne en métabolisme et illustre comment des médicaments ciblés pourraient un jour sauver ou éliminer des protéines spécifiques en les redirigeant dans ou autour de ces circuits de contrôle naturels.
Citation: Andree, G.A., Stier, L.J., Schmiederer, K. et al. Cysteine availability tunes ubiquitin signaling via inverse stability of LRRC58 E3 ligase and its substrate CDO1. Nat Commun 17, 4196 (2026). https://doi.org/10.1038/s41467-026-72524-3
Mots-clés: métabolisme de la cystéine, dégradation des protéines, système ubiquitine, enzyme CDO1, ligase E3