Clear Sky Science · ar
توفر السيستين يضبط إشارات اليوبكويتين عبر استقرار عكسي لبروتين E3 LRRC58 وركيزته CDO1
كيف تقرر الخلايا متى تحفظ الإنزيم أو تتخلص منه
داخل كل خلية، تُبنى جزيئات البروتين وتُهدَم باستمرار. هذا الدفق الصامت يساعد الخلايا على التكيّف مع تغيُّرات الغذاء والضغوط. تكشف هذه الدراسة كيف تستشعر الخلايا البشرية توفر الحمض الأميني المحتوي على الكبريت السيستين ثم تضبط مستويات إنزيم أيضي رئيسي عبر نظام التخلص من البروتينات المصمم بعناية.
مغذٍ يبعث إشارة عندما يكفي الكمية
السيستين هو حمض أميني يجب على الخلايا الحفاظ على توازنه. القِلَّة تهدّد العمليات الحيوية؛ والزيادة قد تكون ضارة. إحدى طرق تعامل الخلايا مع السيستين هي تفكيكه بإنزيم يدعى كايسـتـيين دايوكسيجيناز 1، أو CDO1. أظهرت أعمال سابقة في الحيوانات أنه عندما ينقص السيستين يُدمر CDO1، وعندما يتوافر السيستين يُحفظ CDO1. ومع ذلك، لم يكن واضحًا تمامًا الجهاز الجزيئي الذي يختار CDO1 للإزالة في ظروف نقص السيستين.
العثور على البروتين الذي يعمل كقرص ضبط المغذيات
استخدم الباحثون طريقة مسح بروتينية واسعة النطاق لمسح خلايا بشرية نمت بوجود أو بدون سيستين. ركزوا على عائلة من مجمّعات البروتين تُدعى cullin‑RING ligases، التي تعمل كخطاطيف قابلة للتخصيص لوضع علامة على بروتينات مختارة للتدمير. في الخلايا المحرومة من السيستين، برز مكوّن من هذه الخطاطيف، وهو بروتين مستقبل يُسمى LRRC58: إذ ازداد وجوده وانضم إلى مجمّعات ligase الفعّالة فقط عندما كان السيستين ناقصًا. في الوقت نفسه، انخفضت مستويات CDO1 بشكل حاد، مما يوحي بعلاقة عكسية بين LRRC58 وCDO1. عبر أنواع خلوية متعددة، عندما كانت مستويات LRRC58 مرتفعة في ظل نقص السيستين، كان CDO1 شبه غير قابل للاكتشاف؛ وعند إعادة إضافة السيستين تراجع LRRC58 وعاد CDO1.
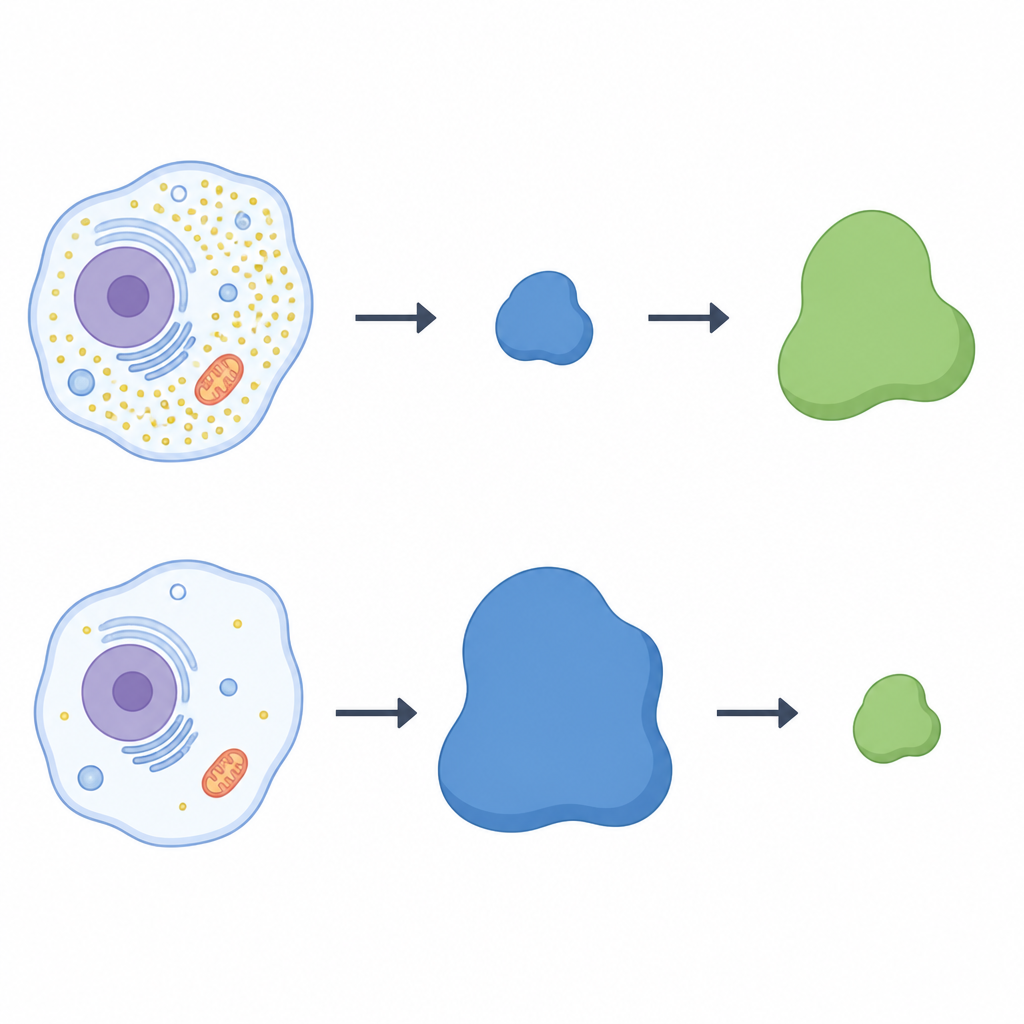
ميزان راقص بين بروتينين
لاختبار ما إذا كان LRRC58 يتحكم مباشرة في CDO1، عطّل الفريق جين LRRC58 في خلايا بشرية. بدون LRRC58، لم يعد CDO1 يختفي أثناء مجاعة السيستين؛ بل استمر وجود الإنزيم حتى عند انخفاض السيستين. إعادة إدخال نسخة فعّالة من LRRC58 أعادت فقدان CDO1، بينما فشلت نسخة متحورة من LRRC58 لا تستطيع الانضمام إلى شركائها في القيام بذلك. أكد نظام مراسل متوهج أن استقرار CDO1 يعتمد على كل من LRRC58 وهياكل cullin‑RING المحددة. مجتمعة، تُظهر هذه التجارب أن LRRC58 وCDO1 يشكّلان زوجًا متقابلاً: LRRC58 عادةً غير مستقر لكنه يستقر عندما ينخفض السيستين، وبدوره يستهدف CDO1 للتدمير في تلك الظروف نفسها.
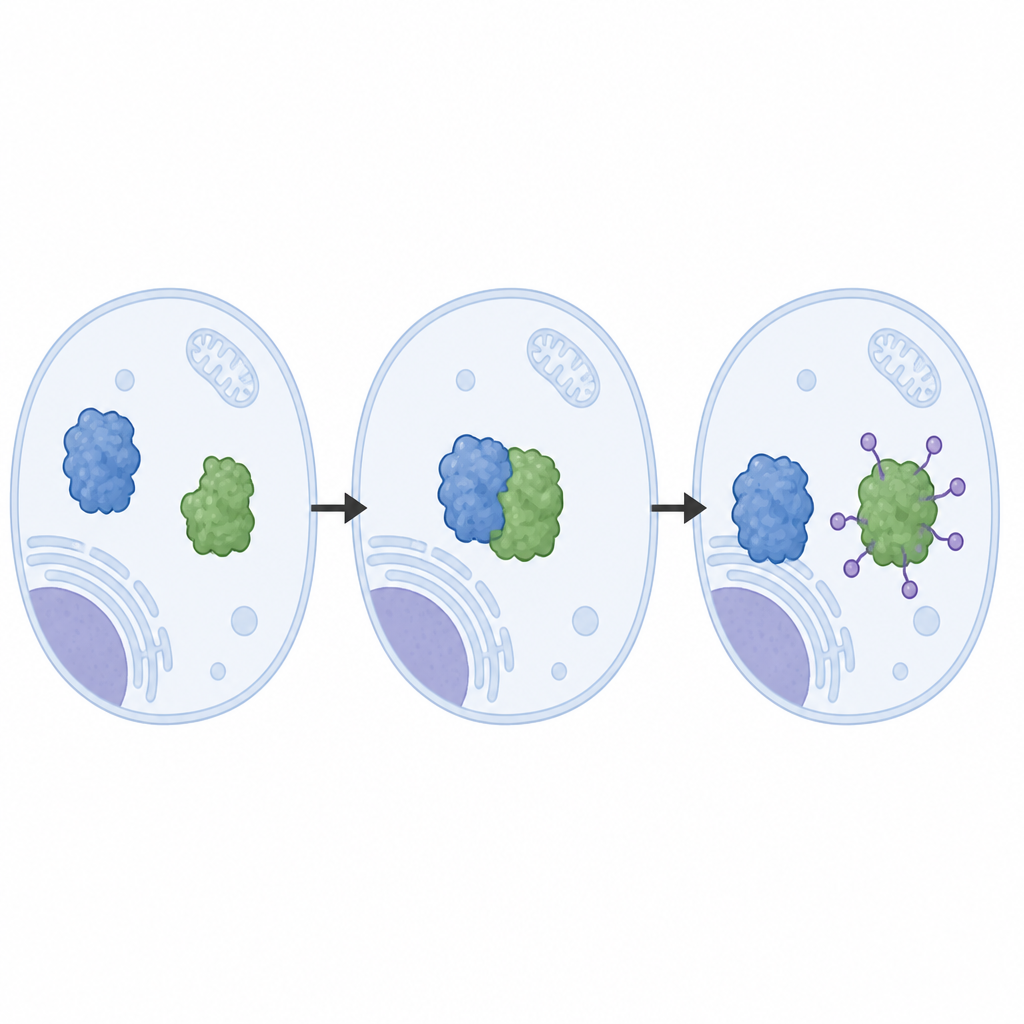
مراقبة آلة الإزالة حتى التفاصيل الذرية
أعاد المؤلفون تركيب نظام LRRC58 من بروتينات منقاة وراقبوا أن LRRC58 يتزاوج مع هياكل قائمة على cullin (CUL2 أو CUL5) ليكوّن آلة وسم فعّالة تُلحِق جزيئات اليوبكويتين بـCDO1. باستخدام المجهر الإلكتروني بالتبريد (cryo‑EM)، حصلوا على صور شبه ذرية لهذه الآلة وهي تعمل. تكشف هذه البنى كيف يمسك LRRC58 بـCDO1 عند عدة نقاط تلامس ويضع حمضًا أمينيًا واحدًا على CDO1، ليسين‑8، بجوار الموقع الكيميائي الذي تُنقل عنده اليوبكويتين. طمس ليسين‑8، أو إضعاف بقع التلامس، يمنع تحلل CDO1 استجابةً لنقص السيستين، مما يؤكد أن الهندسة المرصودة في البنى ضرورية لعمل النظام داخل الخلايا.
التحكم الطبيعي مقابل الاختصارات المحفَّزة دوائيًا
تقارن الدراسة أيضًا هذا المسار الطبيعي مع استراتيجية قائمة على دواء. مركب «الغراء الجزيئي» المطوّر مؤخرًا يفرض التصاق CDO1 بمستقبل مختلف يدعى VHL، الذي يستخدم نفس نوع السقالة القائمة على cullin لكنه يمسك CDO1 بوضعية مغايرة. في الخلايا والتجارب المختبرية، يعلّم هذا الطريق الموجَّه دوائيًا العديد من المواقع على CDO1، وليس فقط ليسين‑8، ويمكنه تدمير CDO1 بفاعلية حتى عندما تعطل طفرات مرتبطة بالمرض تعرفته الطبيعي بواسطة LRRC58. يظهر هذا التباين كيف يمكن للأدوية أن تستغل جهاز التخلص الذاتي داخل الخلية مع تجاوز بعض القيود الطبيعية على مكان وكيفية وسم البروتين.
ماذا يعني هذا لصحة الخلية والعلاجات المستقبلية
لغير المتخصصين، الرسالة الأساسية هي أن الخلايا تستخدم نظامًا مضبوطًا بعناية للتخلص من البروتين لموازنة مستويات الإنزيم مع توافر المغذيات. عندما ينقص السيستين، يستقر LRRC58، يتجمع مع شركائه، ويعلم CDO1 للإزالة انتقائيًا، مما يساعد الخلية على حفظ السيستين. عندما يتوافر السيستين بوفرة، يُحلَّل LRRC58 نفسه، فيُعفى CDO1 حتى يمكن تفكيك السيستين الفائض بأمان. من خلال رسم مسار هذا النظام والتقاطه بدقة عالية، تشرح الدراسة ملاحظة قديمة في الأيض وتوضح كيف قد تُنقِذ الأدوية الموجَّهة أو تزيل بروتينات محددة عبر إعادة توجيهها داخل هذه الدارات التنظيمية الطبيعية أو حولها.
الاستشهاد: Andree, G.A., Stier, L.J., Schmiederer, K. et al. Cysteine availability tunes ubiquitin signaling via inverse stability of LRRC58 E3 ligase and its substrate CDO1. Nat Commun 17, 4196 (2026). https://doi.org/10.1038/s41467-026-72524-3
الكلمات المفتاحية: استقلاب السيستين, تحلل البروتين, نظام اليوبكويتين, إنزيم CDO1, ligase من نوع E3