Clear Sky Science · it
La disponibilità di cisteina regola la segnalazione dell’ubiquitina attraverso la stabilità inversa della ligasi E3 LRRC58 e del suo substrato CDO1
Come le cellule decidono quando salvare o scartare un enzima
All’interno di ogni cellula, le molecole proteiche sono continuamente sintetizzate e degradate. Questo ricambio silenzioso aiuta le cellule ad adattarsi a diete e stress variabili. Questo studio svela come le cellule umane percepiscono la disponibilità dell’amminoacido contenente zolfo cisteina e quindi modulano i livelli di un enzima metabolico chiave, usando un sistema di smaltimento proteico finemente connesso.
Un nutriente che segnala quando è abbastanza
La cisteina è un amminoacido che le cellule devono mantenere in equilibrio. Troppo poca mette a rischio processi vitali; troppo può essere dannosa. Un modo con cui le cellule gestiscono la cisteina è degradarla tramite un enzima chiamato cisteina diossigenasi 1, o CDO1. Studi precedenti negli animali avevano mostrato che quando la cisteina scarseggia, CDO1 viene distrutta, mentre quando la cisteina è abbondante, CDO1 viene preservata. Tuttavia, il macchinario molecolare che seleziona CDO1 per la rimozione in condizioni di carenza di cisteina non era ancora stato identificato chiaramente.
Trovare la proteina che funziona come un regolatore del nutriente
I ricercatori hanno usato un metodo di screening proteico su larga scala per analizzare cellule umane coltivate con o senza cisteina. Si sono concentrati su una famiglia di complessi proteici chiamati ligasi cullina‑RING, che agiscono come ganci personalizzabili che marcano proteine selezionate per la distruzione. Nelle cellule affamate di cisteina, un componente di questi ganci, una proteina recettore chiamata LRRC58, è emersa distintamente: aumentava in abbondanza e si univa ai complessi ligasi attivi solo quando mancava la cisteina. Allo stesso tempo, i livelli di CDO1 precipitavano, suggerendo un legame inverso tra LRRC58 e CDO1. In diversi tipi cellulari, quando LRRC58 era alto in condizioni di bassa cisteina, CDO1 era quasi indetectabile; quando la cisteina veniva reintegrata, LRRC58 diminuiva e CDO1 tornava ai livelli precedenti.
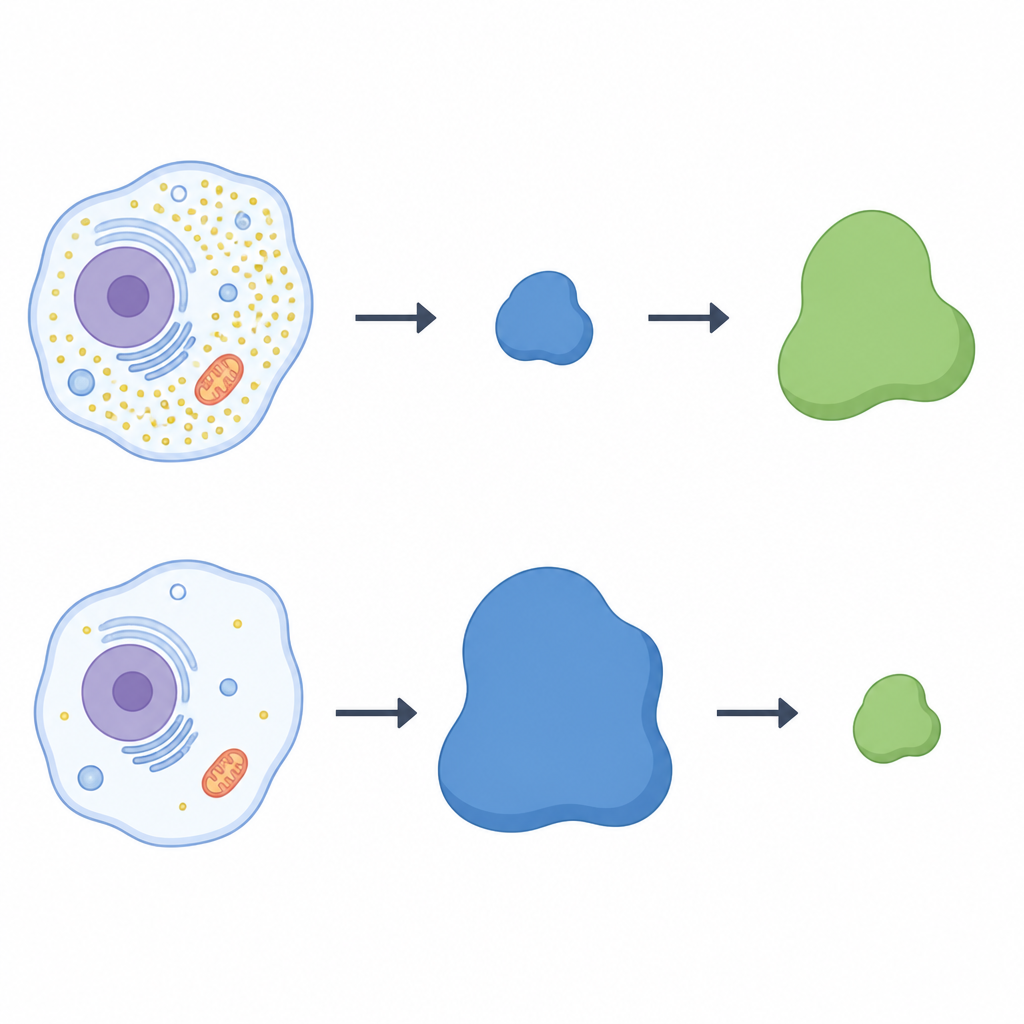
Un bilanciere tra due proteine
Per verificare se LRRC58 controllasse direttamente CDO1, il team ha inattivato il gene LRRC58 in cellule umane. Senza LRRC58, CDO1 non spariva più durante la carenza di cisteina; l’enzima persisteva anche in condizioni di bassa cisteina. Ripristinare una versione funzionale di LRRC58 riportava la perdita di CDO1, mentre una forma mutante di LRRC58 che non può unirsi ai partner della ligasi non riusciva a farlo. Un sistema reporter fluorescente ha confermato che la stabilità di CDO1 dipende sia da LRRC58 sia da specifici impalcature cullina‑RING. Nel loro insieme, questi esperimenti dimostrano che LRRC58 e CDO1 formano una coppia a bilanciere: LRRC58 è normalmente instabile ma si stabilizza quando la cisteina scarseggia, e a sua volta prende di mira CDO1 per la degradazione nelle stesse condizioni.
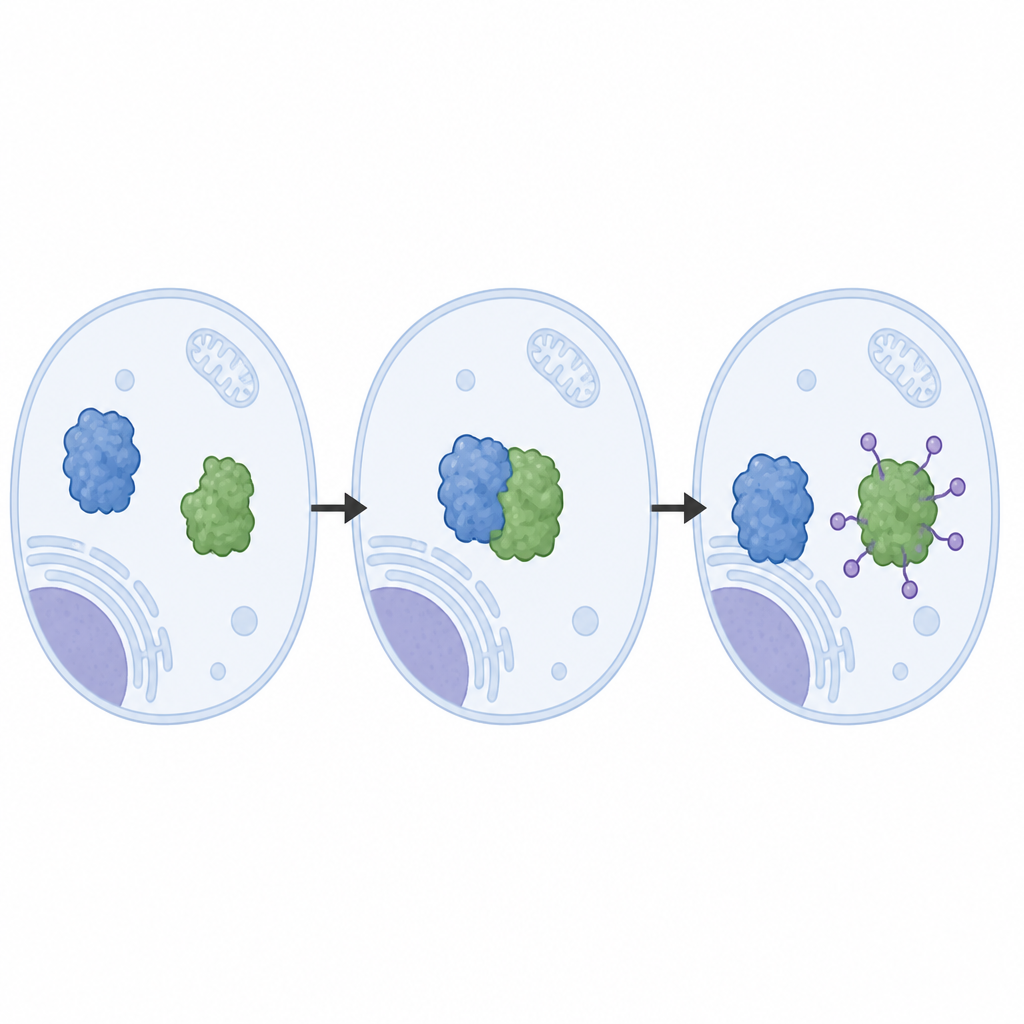
Osservare la macchina di smaltimento a dettaglio atomico
Gli autori hanno ricostruito il sistema LRRC58 a partire da proteine purificate e hanno osservato che LRRC58 si associa con impalcature basate su cullina (CUL2 o CUL5) per formare una macchina attiva che attacca molecole di ubiquitina a CDO1. Usando la crio‑microscopia elettronica, hanno ottenuto immagini quasi atomiche di questa macchina in azione. Queste strutture rivelano come LRRC58 afferra CDO1 in diversi punti di contatto e posiziona un singolo amminoacido di CDO1, la lisina‑8, proprio accanto al sito chimico dove l’ubiquitina viene trasferita. Mutare la lisina‑8, o indebolire i punti di contatto, blocca la degradazione di CDO1 in risposta alla carenza di cisteina, confermando che la geometria vista nelle strutture è essenziale per il funzionamento del sistema nelle cellule.
Controllo naturale versus scorciatoie indotte da farmaci
Lo studio confronta anche questa via naturale con una strategia farmacologica. Un composto sviluppato di recente, una sorta di «collante molecolare», costringe CDO1 ad aderire a un diverso recettore proteico chiamato VHL, che usa lo stesso tipo di impalcatura cullina ma tiene CDO1 in un’orientazione differente. Nelle cellule e nelle reazioni in provetta, questa via guidata dal farmaco marca molti siti su CDO1, non solo la lisina‑8, e può distruggere efficacemente CDO1 anche quando mutazioni legate a malattie compromettono il suo riconoscimento normale da parte di LRRC58. Questo contrasto mostra come i farmaci possano sfruttare l’hardware di smaltimento della cellula pur eludendo alcuni dei vincoli naturali su dove e come una proteina viene marcata.
Cosa significa per la salute cellulare e le future terapie
Per un pubblico non specialista, il messaggio chiave è che le cellule usano un sistema di smaltimento proteico finemente regolato per adattare i livelli enzimatici alla disponibilità di nutrienti. Quando la cisteina scarseggia, LRRC58 si stabilizza, si assemblea con i suoi partner e marca selettivamente CDO1 per la rimozione, aiutando la cellula a conservare cisteina. Quando la cisteina è abbondante, LRRC58 viene esso stesso degradato, risparmiando CDO1 in modo che l’eccesso di cisteina possa essere degradato in sicurezza. Mappando questa via e catturandola ad alta risoluzione, il lavoro spiega un’osservazione di lunga data nel metabolismo e illustra come farmaci mirati potrebbero un giorno salvare o rimuovere proteine specifiche reindirizzandole dentro o attorno a tali circuiti di controllo naturali.
Citazione: Andree, G.A., Stier, L.J., Schmiederer, K. et al. Cysteine availability tunes ubiquitin signaling via inverse stability of LRRC58 E3 ligase and its substrate CDO1. Nat Commun 17, 4196 (2026). https://doi.org/10.1038/s41467-026-72524-3
Parole chiave: metabolismo della cisteina, degradazione proteica, sistema dell’ubiquitina, enzima CDO1, ligasi E3