Clear Sky Science · ru
Доступность цистеина настраивает убиквитин-сигналинг через обратную стабильность E3-лигазы LRRC58 и её субстрата CDO1
Как клетки решают, когда сохранить фермент, а когда его утилизировать
Внутри каждой клетки белки постоянно синтезируются и разрушаются. Эта тихая перестройка помогает клеткам адаптироваться к изменениям питания и стрессам. В работе раскрыто, как человеческие клетки обнаруживают доступность серосодержащей аминокислоты цистеина и точно регулируют уровень ключевого метаболического фермента с помощью тщательно настроенной системы утилизации белков.
Питательное вещество, которое подаёт сигнал «достаточно»
Цистеин — аминокислота, уровень которой клетки должны поддерживать в пределах. Его недостаток угрожает жизненно важным процессам; избыток тоже вреден. Один из способов контролирования цистеина — его разрушение ферментом цистеиндиоксигеназой 1 (CDO1). Ранее в животных моделях показали, что при дефиците цистеина CDO1 разрушается, а при избытке сохраняется. Однако молекулярный аппарат, который выбирает CDO1 для удаления при низком цистеине, долгое время оставался неясным.
Поиск белка, действующего как регулятор уровня питательного вещества
Исследователи использовали масштабный скрининг белков в клетках человека, выращенных с цистеином и без него. Они сосредоточились на семействе комплексов cullin‑RING лигаз, которые действуют как настраиваемые крючки для пометки выбранных белков для разрушения. В клетках, лишённых цистеина, один компонент этих «крючков», рецепторный белок LRRC58, выделялся: он становился более распространённым и включался в активные комплексы лигаз только при отсутствии цистеина. Одновременно уровни CDO1 резко снижались, что указывало на обратную связь между LRRC58 и CDO1. В нескольких типах клеток при высоком уровне LRRC58 в условиях низкого цистеина CDO1 почти не обнаруживался; при добавлении цистеина LRRC58 падал, а CDO1 восстанавливался.
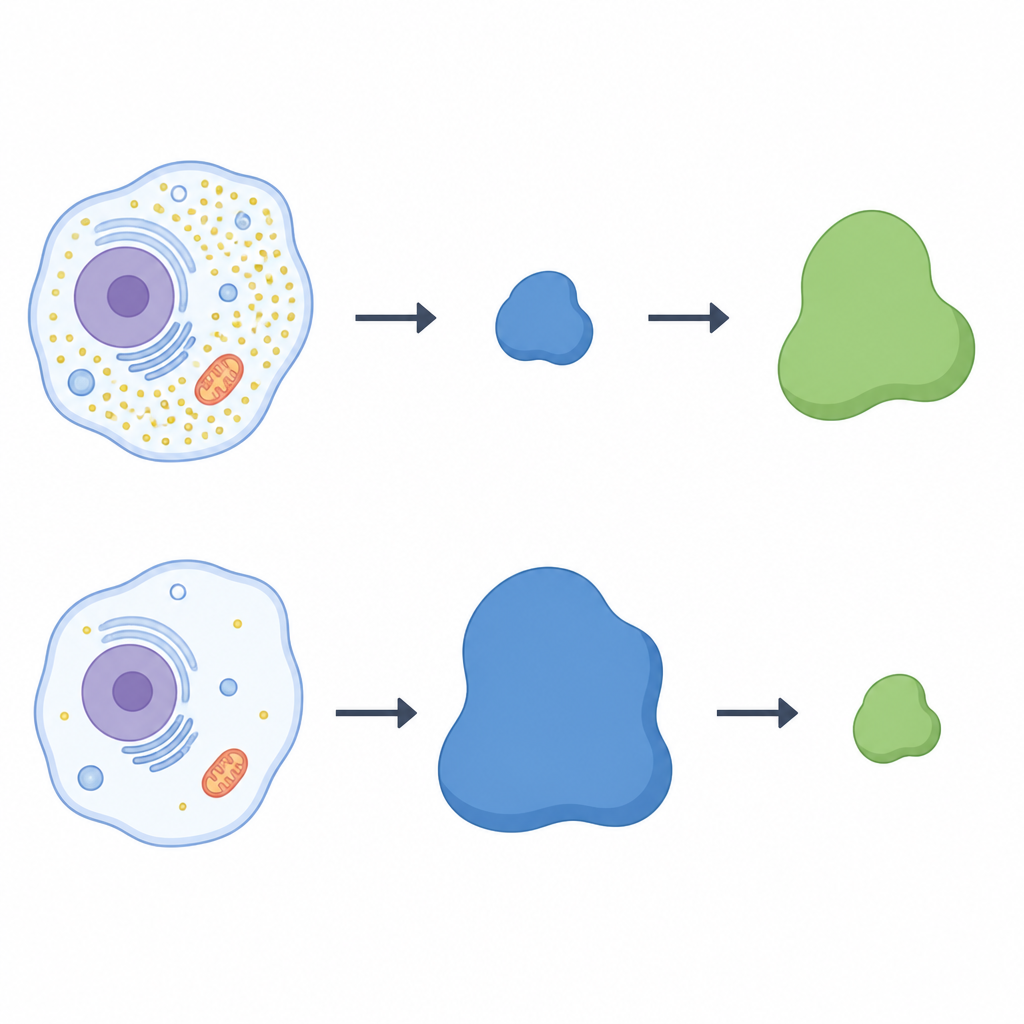
Качели между двумя белками
Чтобы проверить, управляет ли LRRC58 напрямую CDO1, команда инактивировала ген LRRC58 в человеческих клетках. Без LRRC58 CDO1 больше не исчезал при голодании по цистеину; фермент сохранялся даже при низком уровне аминокислоты. Восстановление рабочего варианта LRRC58 вернуло потерю CDO1, тогда как мутант LRRC58, неспособный присоединяться к партнёрам по лигазе, не смог этого сделать. Флуоресцентная репортерная система подтвердила, что стабильность CDO1 зависит как от LRRC58, так и от специфических cullin‑RING каркасов. В совокупности эксперименты демонстрируют, что LRRC58 и CDO1 образуют пару‑качели: LRRC58 обычно нестабилен, но стабилизируется при нехватке цистеина и, в свою очередь, при этих условиях нацеливает CDO1 на уничтожение.
Наблюдение машины утилизации на атомном уровне
Авторы воссоздали систему LRRC58 из очищенных белков и показали, что LRRC58 объединяется с cullin‑скелетами (CUL2 или CUL5), образуя активную машину, которая прикрепляет молекулы убиквитина к CDO1. С помощью криоэлектронной микроскопии они получили изображения машины практически на атомном разрешении. Эти структуры показывают, как LRRC58 захватывает CDO1 в нескольких контактных участках и позиционирует одну аминокислоту на CDO1, лизин‑8, непосредственно рядом с химическим местом передачи убиквитина. Мутация лизина‑8 или ослабление контактных участков блокируют распад CDO1 при дефиците цистеина, подтверждая, что геометрия, видимая в структурах, необходима для работы этой системы в клетках.
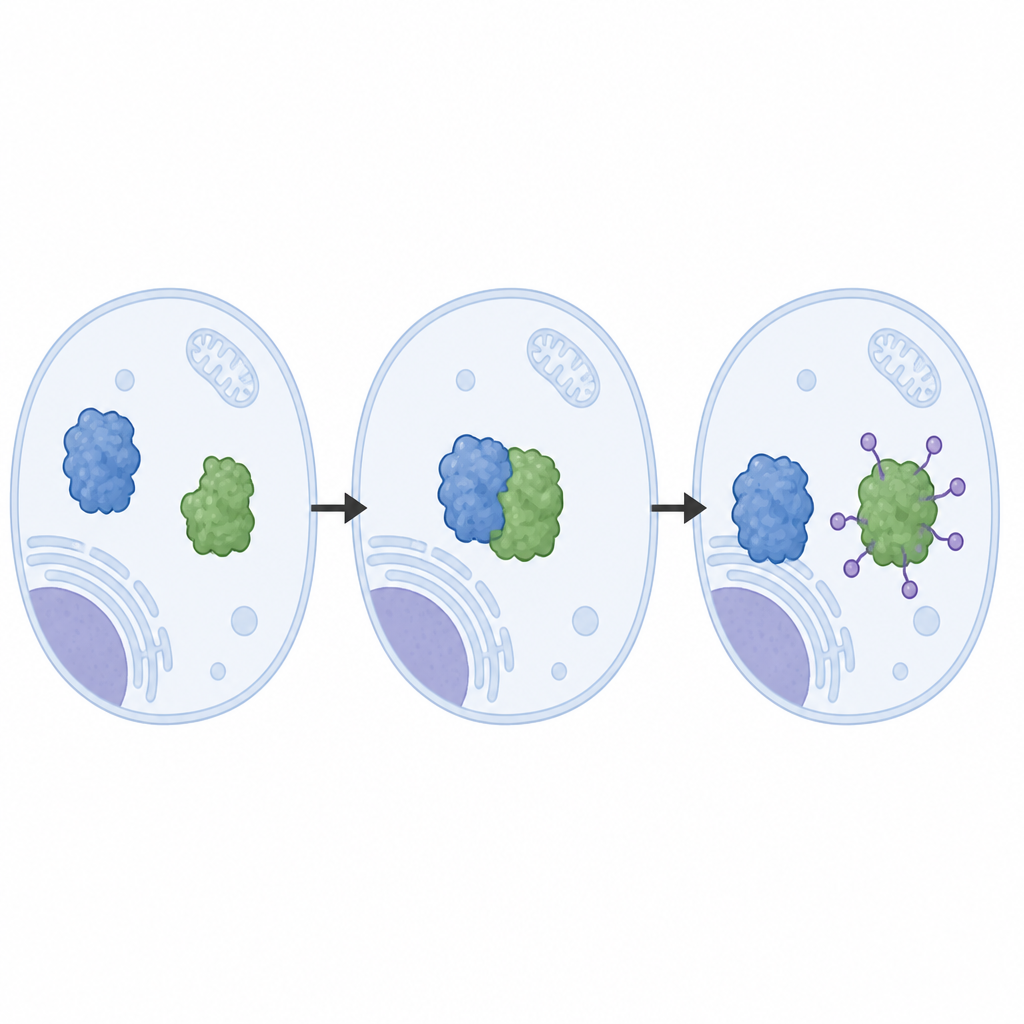
Естественный контроль против лекарственных обходных путей
Исследование также сравнивает этот естественный путь с лекарственной стратегией. Недавно разработанное «молекулярное клейкое» соединение принуждает CDO1 связываться с другим рецептором, VHL, который использует тот же тип cullin‑скелета, но удерживает CDO1 в другой ориентации. В клетках и в пробирке этот направляемый лекарством путь метит множество сайтов на CDO1, а не только лизин‑8, и эффективно уничтожает CDO1 даже когда мутации, связанные с болезнью, нарушают его нормальное распознавание LRRC58. Такое сравнение показывает, как лекарства могут использовать аппарат утилизации клетки, обходя некоторые природные ограничения на то, где и как белок помечается.
Что это значит для здоровья клеток и будущих терапий
Для неспециалиста ключевая мысль такова: клетки используют тонко настроенную систему утилизации белков, чтобы согласовать уровни ферментов с доступностью питательных веществ. Когда цистеина мало, LRRC58 стабилизируется, собирается с партнёрами и избирательно помечает CDO1 для удаления, помогая клетке экономить цистеин. Когда цистеина достаточно, сам LRRC58 разрушается, щадя CDO1, чтобы избыток цистеина мог быть безопасно разложен. Карта этого пути и его получение в высоком разрешении объясняют давнее наблюдение в метаболизме и иллюстрируют, как таргетные препараты однажды могут спасать или удалять конкретные белки, перенаправляя их внутрь или вокруг таких природных контролирующих цепочек.
Цитирование: Andree, G.A., Stier, L.J., Schmiederer, K. et al. Cysteine availability tunes ubiquitin signaling via inverse stability of LRRC58 E3 ligase and its substrate CDO1. Nat Commun 17, 4196 (2026). https://doi.org/10.1038/s41467-026-72524-3
Ключевые слова: метаболизм цистеина, деградация белков, система убиквитина, фермент CDO1, E3-лигаза