Clear Sky Science · es
La disponibilidad de cisteína ajusta la señalización por ubiquitina mediante la estabilidad inversa de la ligasa E3 LRRC58 y su sustrato CDO1
Cómo deciden las células cuándo salvar o desechar una enzima
Dentro de cada célula, las moléculas proteicas se sintetizan y se degradan constantemente. Este silencioso reciclaje ayuda a las células a adaptarse a cambios en la dieta y a situaciones de estrés. Este estudio revela cómo las células humanas detectan la disponibilidad del aminoácido que contiene azufre, la cisteína, y ajustan con precisión los niveles de una enzima metabólica clave mediante un sistema de eliminación de proteínas cuidadosamente cableado.
Un nutriente que indica cuándo hay suficiente
La cisteína es un aminoácido que las células deben mantener en equilibrio. Su defecto amenaza procesos vitales; su exceso puede ser dañino. Una manera de gestionar la cisteína es degradarla con una enzima llamada cisteína dioxigenasa 1, o CDO1. Estudios previos en animales mostraron que cuando la cisteína escasea, CDO1 se destruye, y cuando la cisteína abunda, CDO1 se preserva. Sin embargo, la maquinaria molecular que selecciona a CDO1 para su eliminación en condiciones de baja cisteína no se había identificado con claridad.
Encontrar la proteína que actúa como un regulador de nutrientes
Los investigadores emplearon un método de cribado proteico a gran escala para analizar células humanas cultivadas con o sin cisteína. Se centraron en una familia de complejos proteicos llamados ligasas cullina‑RING, que actúan como ganchos personalizables que etiquetan proteínas seleccionadas para su destrucción. En las células privadas de cisteína, un componente de estos ganchos, una proteína receptora llamada LRRC58, destacó: aumentó su abundancia y se incorporó a complejos de ligasa activos únicamente cuando faltaba cisteína. Al mismo tiempo, los niveles de CDO1 cayeron drásticamente, lo que sugiere un vínculo inverso entre LRRC58 y CDO1. En varios tipos celulares, cuando LRRC58 era alto en condiciones de baja cisteína, CDO1 era prácticamente indetectable; al reponer cisteína, LRRC58 disminuía y CDO1 se recuperaba.
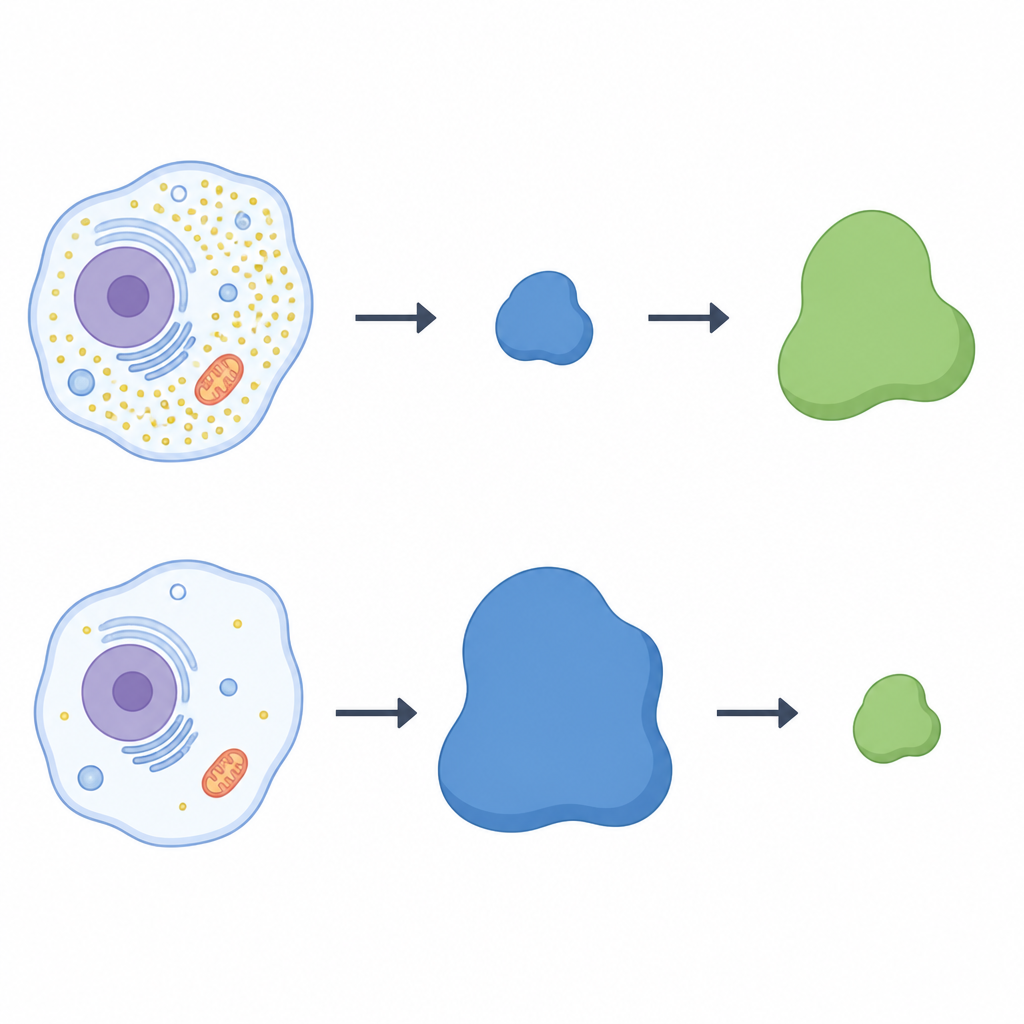
Un balancín entre dos proteínas
Para comprobar si LRRC58 controla directamente a CDO1, el equipo desactivó el gen LRRC58 en células humanas. Sin LRRC58, CDO1 dejó de desaparecer durante la privación de cisteína; la enzima persistió incluso con bajos niveles de cisteína. Reintroducir una versión funcional de LRRC58 restauró la pérdida de CDO1, mientras que una forma mutante de LRRC58 que no puede asociarse con sus socios de la ligasa no lo logró. Un sistema reportero fluorescente confirmó que la estabilidad de CDO1 depende tanto de LRRC58 como de andamios cullin‑RING específicos. En conjunto, estos experimentos muestran que LRRC58 y CDO1 forman un par tipo balancín: LRRC58 es normalmente inestable pero se estabiliza cuando la cisteína escasea, y a su vez dirige a CDO1 hacia la destrucción en esas mismas condiciones.
Observar la máquina de eliminación a detalle atómico
Los autores reconstruyeron el sistema LRRC58 a partir de proteínas purificadas y observaron que LRRC58 se asocia con andamios basados en cullina (CUL2 o CUL5) para formar una máquina de etiquetado activa que añade moléculas de ubiquitina a CDO1. Mediante criomicroscopía electrónica obtuvieron imágenes casi atómicas de esta máquina en acción. Estas estructuras revelan cómo LRRC58 sujeta a CDO1 en varios puntos de contacto y posiciona un único aminoácido de CDO1, la lisina‑8, justo junto al sitio químico donde se transfiere la ubiquitina. Mutar la lisina‑8, o debilitar los parches de contacto, impide la degradación de CDO1 en respuesta a la escasez de cisteína, lo que confirma que la geometría observada en las estructuras es esencial para el funcionamiento del sistema dentro de las células.
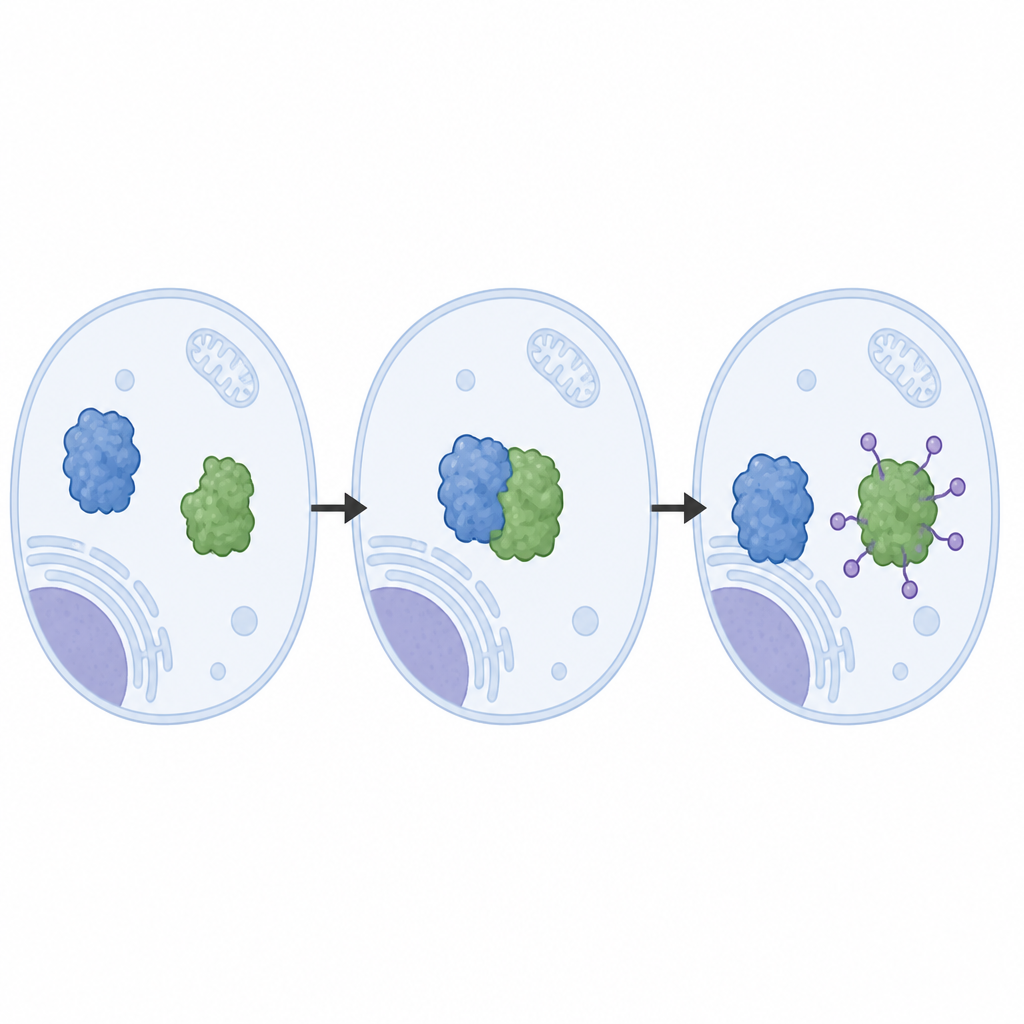
Control natural frente a atajos inducidos por fármacos
El estudio también compara esta vía natural con una estrategia basada en fármacos. Un compuesto tipo “pegamento molecular” desarrollado recientemente fuerza que CDO1 se adhiera a otra proteína receptora llamada VHL, que utiliza el mismo tipo de andamio cullin pero mantiene a CDO1 en una orientación diferente. En células y reacciones in vitro, esta ruta guiada por el fármaco marca muchos sitios en CDO1, no solo la lisina‑8, y puede destruir eficientemente CDO1 incluso cuando mutaciones vinculadas a enfermedad alteran su reconocimiento normal por LRRC58. Este contraste muestra cómo los fármacos pueden aprovechar el propio hardware de eliminación celular al tiempo que evitan algunas de las limitaciones naturales sobre dónde y cómo se etiqueta una proteína.
Qué significa esto para la salud celular y futuras terapias
Para un público no especialista, el mensaje clave es que las células usan un sistema de eliminación de proteínas finamente ajustado para adaptar los niveles enzimáticos al suministro de nutrientes. Cuando la cisteína escasea, LRRC58 se estabiliza, se ensambla con sus socios y marca selectivamente a CDO1 para su eliminación, ayudando a la célula a conservar cisteína. Cuando la cisteína abunda, LRRC58 se degrada, lo que preserva a CDO1 para que el exceso de cisteína pueda ser descompuesto de forma segura. Al cartografiar esta vía y captar su estructura a alta resolución, el trabajo explica una observación de larga data en metabolismo e ilustra cómo fármacos dirigidos podrían algún día rescatar o eliminar proteínas específicas redirigiéndolas hacia —o alrededor de— estos circuitos de control natural.
Cita: Andree, G.A., Stier, L.J., Schmiederer, K. et al. Cysteine availability tunes ubiquitin signaling via inverse stability of LRRC58 E3 ligase and its substrate CDO1. Nat Commun 17, 4196 (2026). https://doi.org/10.1038/s41467-026-72524-3
Palabras clave: metabolismo de la cisteína, degradación de proteínas, sistema de ubiquitina, enzima CDO1, ligasa E3