Clear Sky Science · pl
Dostępność cysteiny reguluje sygnalizację ubikwitynową poprzez odwrotną stabilność ligazy E3 LRRC58 i jej substratu CDO1
Jak komórki decydują, kiedy zachować, a kiedy złomować enzym
W każdej komórce białka są nieustannie syntetyzowane i rozkładane. Ten cichy ruchomek pomaga komórkom dostosowywać się do zmieniającej się diety i stresów. W pracy tej odkryto, jak komórki ludzkie wykrywają dostępność siarko‑zawierającego aminokwasu cysteiny i precyzyjnie regulują poziomy kluczowego enzymu metabolicznego, wykorzystując starannie zaprojektowany system usuwania białek.
Składnik odżywczy, który sygnalizuje: dość
Cysteina to aminokwas, którego komórki muszą utrzymywać w równowadze. Zbyt mało zagraża podstawowym procesom; zbyt dużo może być szkodliwe. Jednym ze sposobów kontrolowania cysteiny jest jej rozkład przez enzym cysteiny dioksygenazę 1, czyli CDO1. W badaniach na zwierzętach wykazano wcześniej, że gdy cysteiny brakuje, CDO1 ulega zniszczeniu, a gdy cysteiny jest pod dostatkiem, CDO1 jest zachowywany. Jednak molekularne mechanizmy wybierające CDO1 do usunięcia w warunkach niedoboru cysteiny nie były wcześniej jasno zidentyfikowane.
Poszukiwanie białka działającego jak regulator odżywienia
Naukowcy zastosowali szeroko zakrojone przesiewy białkowe na komórkach ludzkich hodowanych z cysteiną lub bez niej. Skoncentrowali się na rodzinie kompleksów zwanych ligazami cullin‑RING, które działają jak konfigurowalne haki znakujące wybrane białka do degradacji. W komórkach pozbawionych cysteiny jeden z komponentów haka, receptor o nazwie LRRC58, wyróżniał się: stawał się bardziej obfity i włączał się do aktywnych kompleksów ligaz tylko wtedy, gdy brakowało cysteiny. Jednocześnie poziomy CDO1 gwałtownie spadały, co sugerowało odwrotną zależność między LRRC58 a CDO1. W różnych typach komórek, gdy LRRC58 był wysoki przy niskiej cysteinie, CDO1 był prawie niewykrywalny; po przywróceniu cysteiny LRRC58 malał, a CDO1 wracał.
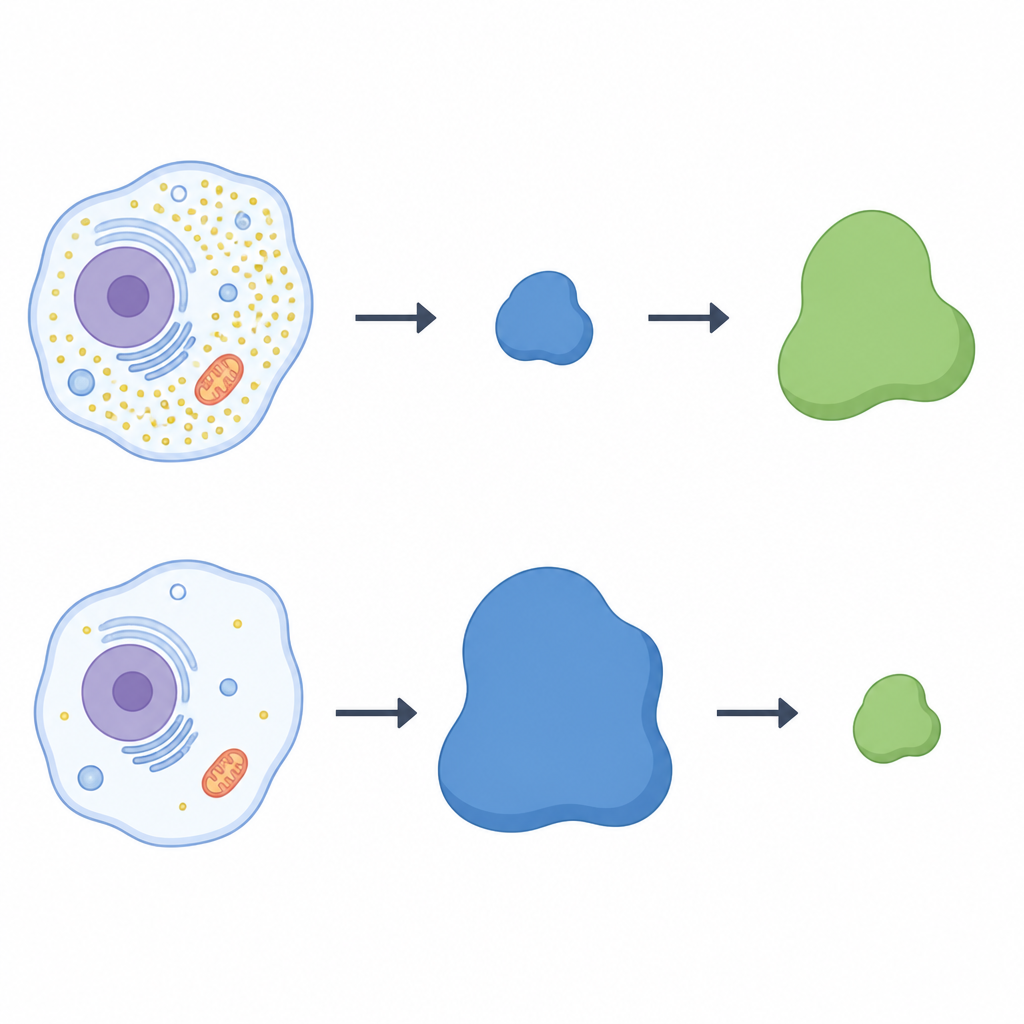
Huśtawka między dwoma białkami
Aby sprawdzić, czy LRRC58 bezpośrednio kontroluje CDO1, zespół wyłączył gen LRRC58 w komórkach ludzkich. Bez LRRC58 CDO1 przestał zanikać podczas głodzenia w cysteinie; enzym utrzymywał się nawet przy niskim stężeniu cysteiny. Przywrócenie funkcjonalnej wersji LRRC58 odtworzyło utratę CDO1, podczas gdy mutant LRRC58, który nie potrafi włączyć się do partnerów ligazowych, nie zdołał tego zrobić. System z fluorescencyjnym reporterem potwierdził, że stabilność CDO1 zależy zarówno od LRRC58, jak i od konkretnych rusztowań cullin‑RING. Razem te eksperymenty pokazują, że LRRC58 i CDO1 tworzą parę huśtawek: LRRC58 zwykle jest nietrwały, lecz stabilizuje się, gdy cysteiny brakuje, i wówczas oznacza CDO1 do zniszczenia w tych samych warunkach.
Obserwowanie maszyny do usuwania na poziomie atomowym
Autorzy odtworzyli system LRRC58 z oczyszczonych białek i zaobserwowali, że LRRC58 łączy się z rusztowaniami opartymi na cullinach (CUL2 lub CUL5), tworząc aktywną maszynę znakującą, która przyłącza cząsteczki ubikwityny do CDO1. Z wykorzystaniem krio‑elektronowej mikroskopii uzyskali obrazy tej maszyny niemal w skali atomowej. Struktury te pokazują, jak LRRC58 chwyta CDO1 w kilku punktach styku i pozycjonuje pojedynczy resztę aminokwasową na CDO1 — lizynę‑8 — tuż obok miejsca chemicznego, gdzie przekazywana jest ubikwityna. Mutacje lizyny‑8 lub osłabienie punktów styku blokują rozkład CDO1 w reakcji na niedobór cysteiny, potwierdzając, że geometria widoczna w strukturach jest niezbędna dla funkcjonowania systemu w komórkach.
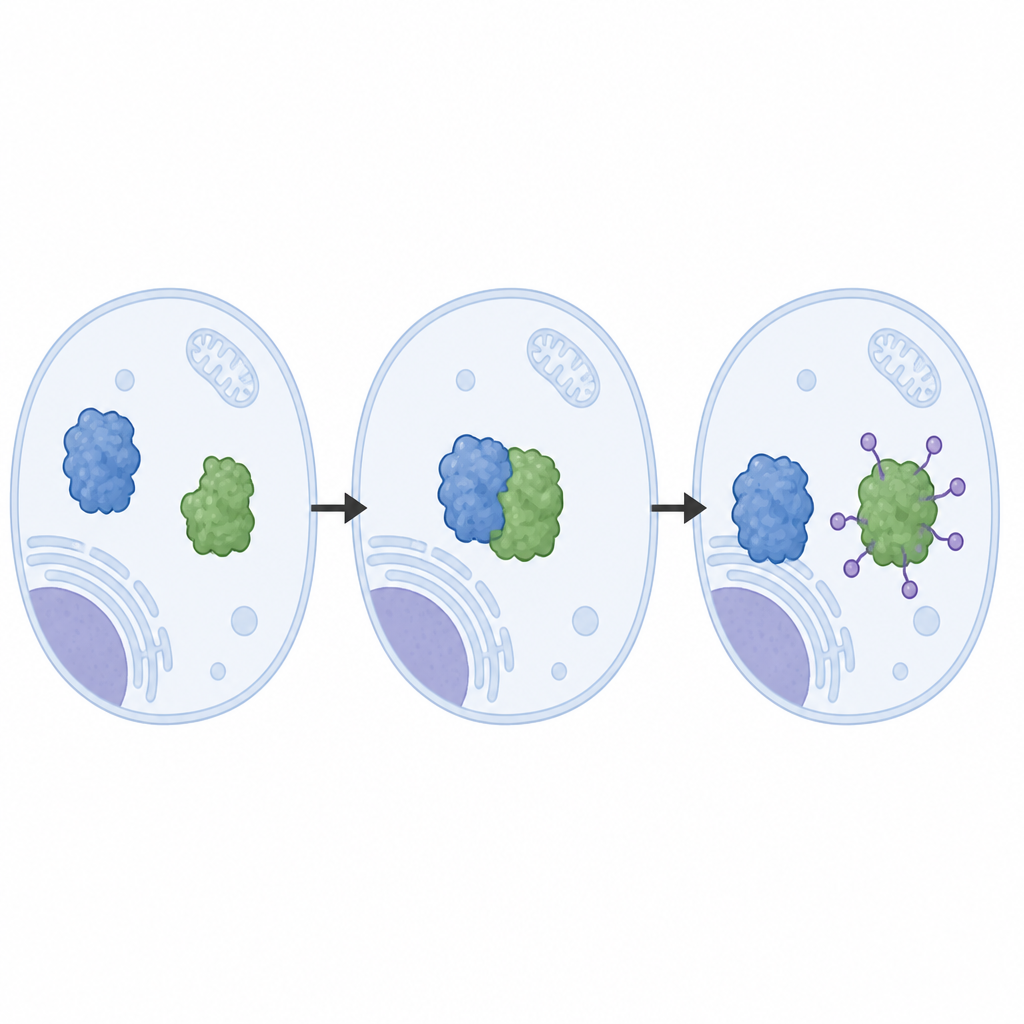
Naturalna kontrola kontra farmakologiczne skróty
Badanie porównuje też tę naturalną ścieżkę z podejściem opartym na lekach. Niedawno opracowany związek typu „molecular glue” zmusza CDO1 do przyłączenia się do innego receptora zwanego VHL, który używa tego samego typu rusztowania cullinowego, lecz trzyma CDO1 w innej orientacji. W komórkach i reakcjach in vitro ta naprowadzona przez lek droga znakuję wiele miejsc na CDO1, nie tylko lizynę‑8, i może skutecznie zniszczyć CDO1 nawet wtedy, gdy mutacje związane z chorobami zaburzają jego normalne rozpoznanie przez LRRC58. Kontrast ten pokazuje, jak leki mogą wykorzystać sprzęt komórkowy do usuwania białek, jednocześnie omijając niektóre naturalne ograniczenia dotyczące miejsca i sposobu znakowania białka.
Co to oznacza dla zdrowia komórek i przyszłych terapii
Dla czytelnika niebędącego specjalistą główne przesłanie jest takie, że komórki używają precyzyjnie wyregulowanego systemu usuwania białek, aby dopasować poziomy enzymów do dostępności składników odżywczych. Gdy cysteiny brakuje, LRRC58 stabilizuje się, łączy z partnerami i selektywnie oznacza CDO1 do usunięcia, pomagając komórce oszczędzać cysteinę. Gdy cysteiny jest dużo, LRRC58 ulega degradacji, oszczędzając CDO1, aby nadmiar cysteiny mógł zostać bezpiecznie rozłożony. Mapując tę ścieżkę i rejestrując ją w wysokiej rozdzielczości, praca wyjaśnia długo obserwowane zjawisko w metabolizmie i ilustruje, jak ukierunkowane leki mogłyby w przyszłości ratować lub usuwać określone białka, kierując je w obrębie lub wokół takich naturalnych obwodów kontrolnych.
Cytowanie: Andree, G.A., Stier, L.J., Schmiederer, K. et al. Cysteine availability tunes ubiquitin signaling via inverse stability of LRRC58 E3 ligase and its substrate CDO1. Nat Commun 17, 4196 (2026). https://doi.org/10.1038/s41467-026-72524-3
Słowa kluczowe: metabolizm cysteiny, degradacja białek, układ ubikwitynowy, enzym CDO1, ligaza E3