Clear Sky Science · zh
不同界面脂质对糖类转运蛋白功能性二聚化的多样调控
细胞脂肪如何操控微小的糖类通道
我们的细胞充满了微观的守门者,它们在油性膜间搬运糖类和其他养分。这项研究提出了一个看似简单但影响深远的问题:周围的脂肪分子本身能否决定某个糖类通道是否形成正确的构象并发挥功能?通过在近乎天然的膜环境中观察一种细菌糖转运蛋白,作者们表明少数特定的脂质在恰当位置的存在,能够左右通道是顺畅工作还是发生卡滞。
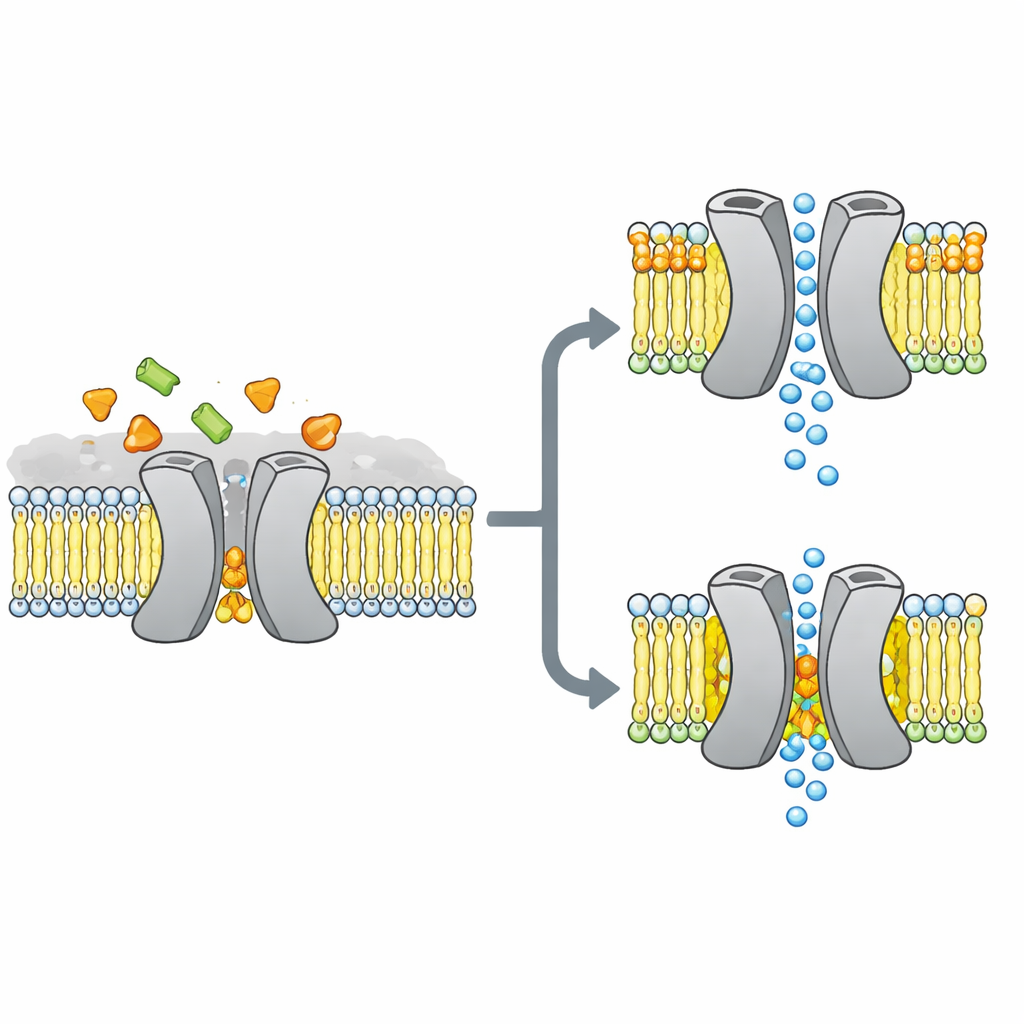
故事核心的糖类通道
研究者关注的是VsSemiSWEET,一种来自细菌的糖类转运蛋白,与植物和动物中的同类蛋白为近亲。这些蛋白跨越细胞膜,通常以一对(二聚体)形式工作,让糖类从一侧通过到另一侧。早期的晶体结构揭示了这些二聚体的总体形状,但奇怪的是未见结合的脂质,尽管其他方法提示某些脂质会紧密依附于转运蛋白并可能控制其行为。
寻找隐秘的脂质伙伴
为了在更接近生理的环境中识别哪些脂质真正黏附于该转运蛋白,团队将VsSemiSWEET重新组装进由细菌脂质构成的膜片,并使用固态核磁共振(一种能在完整膜中研究蛋白的强大磁共振方法)。他们检测到三类共洗脱的脂质:磷脂酰乙醇胺(PE)、磷脂酰甘油(PG)和心磷脂(CDL)。这三种脂质都位于两个蛋白伙伴的接触面处,但心磷脂结合得尤其牢固,能在去除大多数其他脂质的强力去污剂处理下仍然保留。
在原生栖息地中观察二聚体
通过使用精心标记的样品与多维核磁实验的组合,作者建立了嵌入脂质双层中的转运蛋白二聚体的高分辨率三维结构。他们证实蛋白确实形成配对结构,朝膜外侧开放的腔体适合糖类进入。当他们在二聚体界面上对接触脂质的三个带正电氨基酸进行突变时,配对结构变得脆弱:在凝胶中更容易解体,核磁信号展宽并消失,表明复合体变得柔性且不稳定。计算机模拟也反映了这一情形,显示在缺乏界面脂质的情况下,两侧伙伴会分离并失去许多稳定接触点。
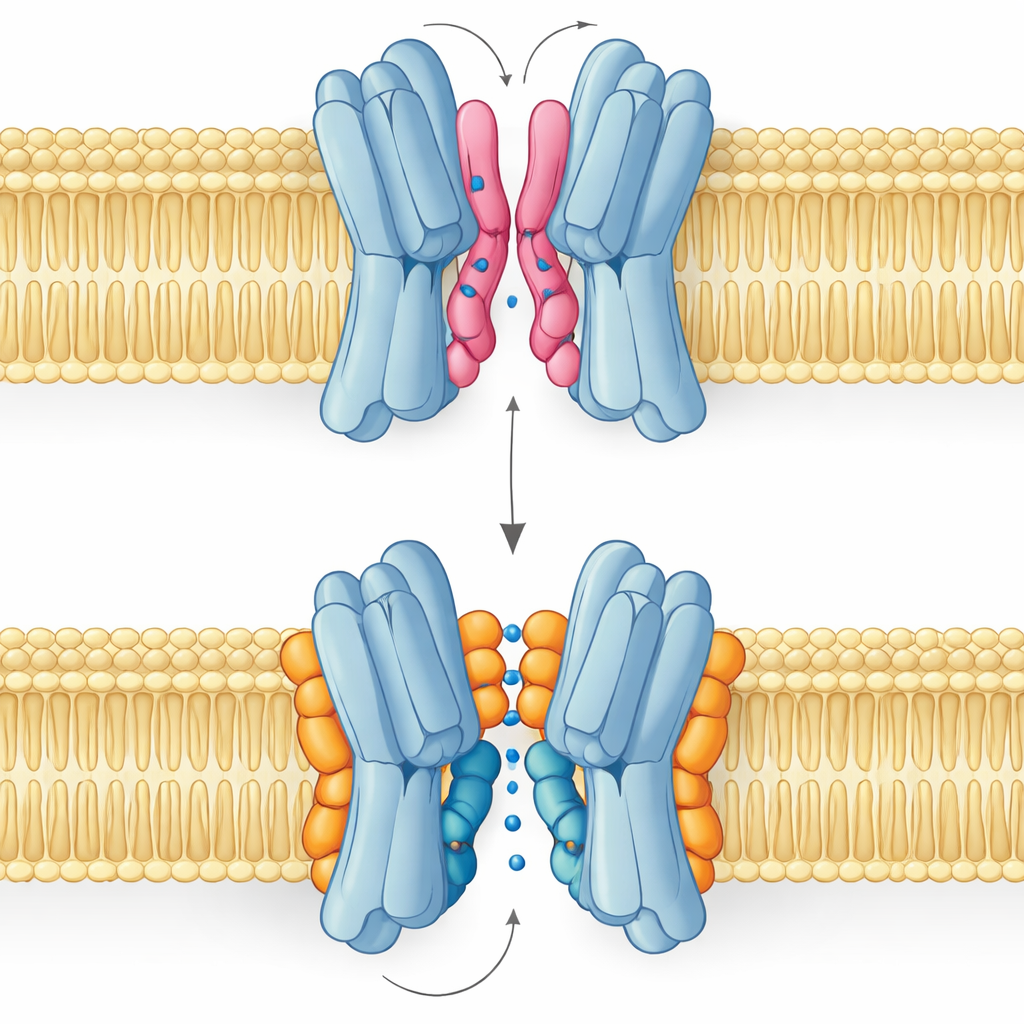
良脂与劣脂:两种结合模式的故事
最令人惊讶的结果是,并非所有界面脂质都以相同方式提供帮助。模拟与核磁测量显示,PG 与心磷脂在重叠位点结合,但采用不同的构象。带两个尾链且带单负电荷的PG可以将其尾链深入蛋白伙伴之间狭窄的缝隙,略微撬开它们,甚至堵塞中央通道。相比之下,心磷脂头部体积更大,带双负电荷并拥有四条尾链。它的头部像夹具一样抓住一簇碱性残基,而更庞大的尾束则停留在外表面,避免进入中央腔体,转而支撑在运输周期中发生摆动的关键螺旋之一。
从脂质结合到糖类转运
这些结构差异直接转化为功能。来自长时程模拟的自由能景观显示,当心磷脂结合时,转运蛋白可以顺利访问外向开放、闭塞和内向开放等完成糖类穿膜所需的构象。最稳定的状态是面向外侧,与实验结构一致,但通向内向构象的路径仍可达成。在PG主导的情况下,情形改变:其尾链填满中央腔体,亚基间的接触变得紊乱,蛋白被困在一种非生产性的构象中,糖类无法通过。PE 的行为与 PG 类似,尽管它促成亚基配对,但使它们比心磷脂结合时更为异质且不那么稳定。
为何这超越了单一细菌转运蛋白的重要性
通俗地说,这项工作表明膜脂并非只是被动的润滑物,而是能附着于蛋白、支撑可动部件或楔入关键缝隙的主动机械元件。对于VsSemiSWEET,心磷脂像量身定制的垫片,一方面锁定两个半体,另一方面引导完成糖转运所需的运动,而PG则倾向于使系统卡住。由于许多膜蛋白具有相似的界面脂质位点并常依赖心磷脂或相关脂质,这些发现为如何通过改变脂质组成来调节转运蛋白、通道和泵的活性提供了更广泛的蓝图。
引用: Zhang, Y., Zhao, W., Duan, M. et al. Diverse regulation of functional dimerization of a sugar transporter by different interfacial lipids. Nat Commun 17, 3062 (2026). https://doi.org/10.1038/s41467-026-69804-3
关键词: 膜脂质, 糖类转运蛋白, 心磷脂, 蛋白质二聚化, 固态核磁共振