Clear Sky Science · it
Regolazione diversificata della dimerizzazione funzionale di un trasportatore di zuccheri da parte di diversi lipidi interfaciali
Come i grassi cellulari guidano piccoli cancelli per zuccheri
Le nostre cellule sono piene di guardiani microscopici che spostano zuccheri e altri nutrienti attraverso membrane oleose. Questo studio pone una domanda apparentemente semplice ma dalle grandi implicazioni: i grassi circostanti possono decidere da soli se uno di questi cancelli per zuccheri assuma la forma corretta e funzioni davvero? Osservando un trasportatore di zuccheri batterico in una membrana quasi naturale, gli autori mostrano che poche molecole lipidiche specifiche, posizionate nel punto giusto, possono fare la differenza tra un cancello che funziona regolarmente e uno inceppato.
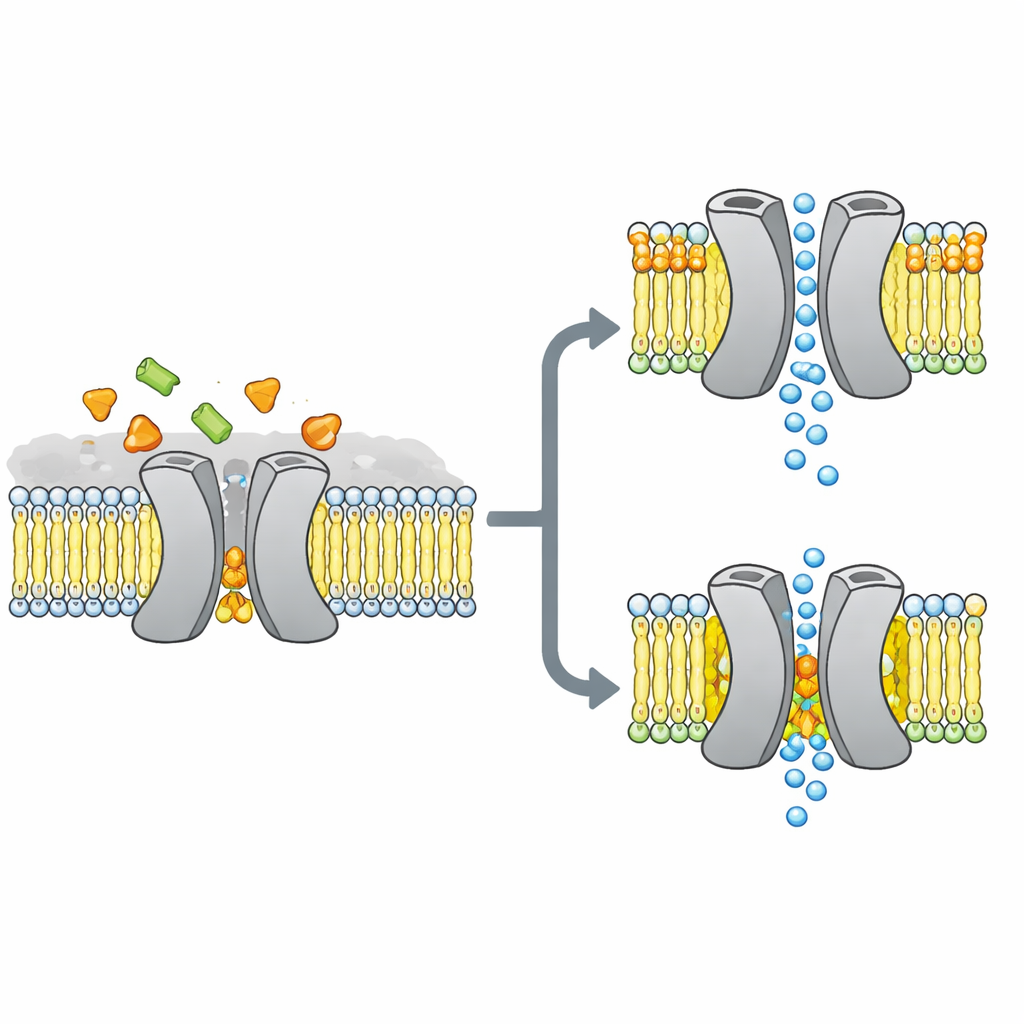
Il cancello per zuccheri al centro della storia
I ricercatori si sono concentrati su VsSemiSWEET, un parente batterico dei trasportatori di zuccheri presenti in piante e animali. Queste proteine attraversano la membrana cellulare e normalmente lavorano in coppia, o dimeri, per consentire allo zucchero di passare da un lato all’altro. Strutture cristallografiche precedenti avevano mostrato la forma complessiva di queste coppie ma, curiosamente, non evidenziavano lipidi legati, nonostante altri metodi suggerissero che certi lipidi si attaccano saldamente al trasportatore e potrebbero controllarne il comportamento.
Alla ricerca dei partner lipidici nascosti
Per scoprire quali lipidi si legano realmente a questo trasportatore in un contesto verosimile, il gruppo ha ricostituito VsSemiSWEET in frammenti di membrana preparati con lipidi batterici e ha impiegato la NMR in stato solido, una potente forma di risonanza magnetica che funziona su proteine all’interno di membrane integre. Hanno rilevato tre tipi di lipidi co‑purificanti: fosfatidiletanolamina (PE), fosfatidilglicerolo (PG) e cardiolipina (CDL). Tutti e tre si annidano nella superficie di contatto tra i due partner proteici, ma la cardiolipina si lega in modo particolarmente saldo, resistendo a lavaggi con detergenti aggressivi che asportano la maggior parte degli altri lipidi.
Vedere il dimero nel suo habitat nativo
Usando una combinazione di campioni opportunamente marcati ed esperimenti NMR multidimensionali, gli autori hanno costruito una struttura 3D ad alta risoluzione del dimero del trasportatore all’interno di un doppio strato lipidico. Hanno confermato che la proteina forma effettivamente un assemblaggio a coppia con una cavità aperta verso l’esterno della cellula, adatta all’ingresso dello zucchero. Quando hanno mutato tre aminoacidi carichi positivamente all’interfaccia del dimero che entrano in contatto con i lipidi, la struttura a coppia è diventata fragile: nei gel si disfaceva più facilmente e i suoi segnali NMR si allargavano e scomparivano, indicando un complesso flessibile e instabile. Simulazioni al computer hanno confermato questo quadro, mostrando che senza lipidi interfaciali i due partner si allontanano e perdono molti contatti stabilizzanti.
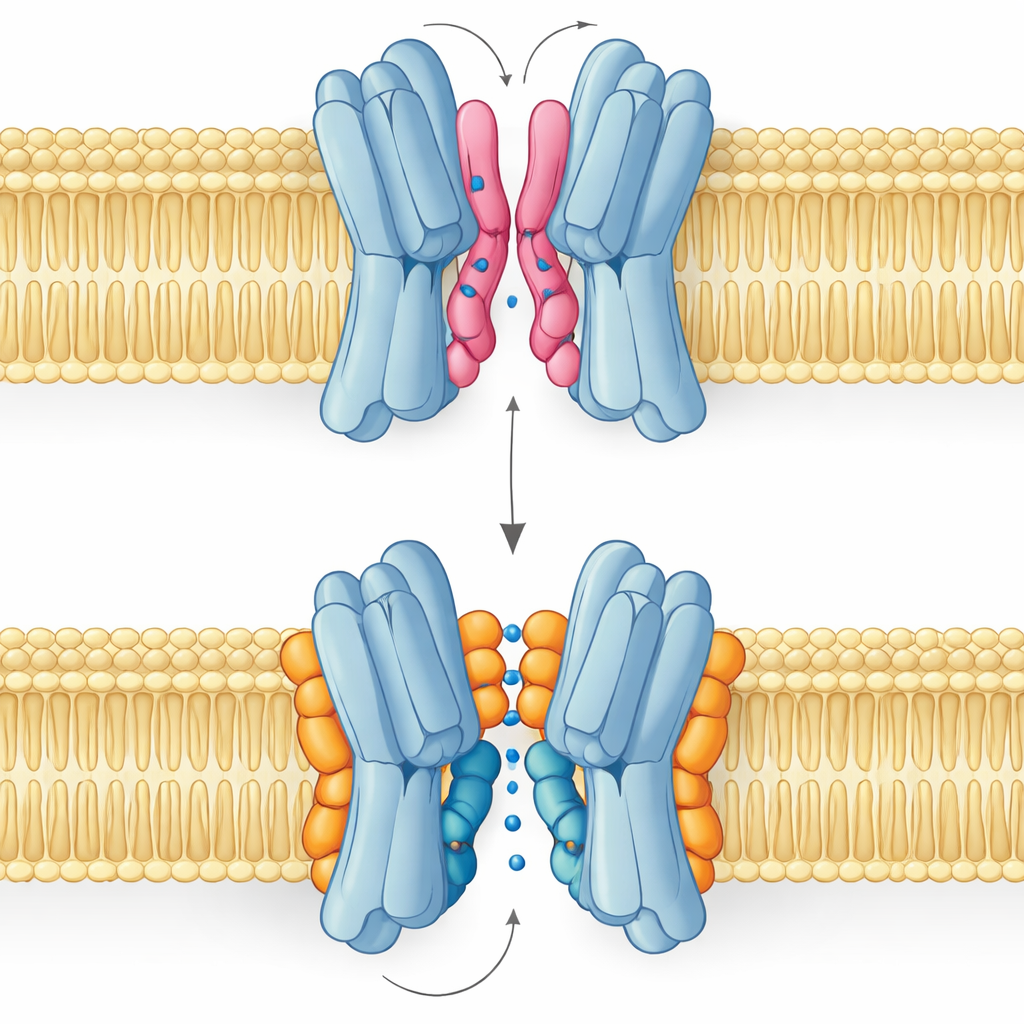
Buon lipide, lipide dannoso: la storia di due modalità di legame
Il risultato più sorprendente è che non tutti i lipidi di interfaccia aiutano allo stesso modo. Simulazioni e misure NMR hanno rivelato che PG e cardiolipina si legano in punti sovrapposti ma assumono pose distinte. Il PG, con due code e una singola carica negativa, può insinuare le sue code in profondità nell’angusto spazio tra i partner proteici, separandoli leggermente e persino occludendo il passaggio centrale. La cardiolipina, al contrario, ha una testa più ingombrante con due cariche negative e quattro code. La sua testa afferra un cluster di residui basici come una pinza, mentre il più voluminoso fascio di code resta sulla superficie esterna, evitando la cavità centrale e invece rinforzando una delle eliche chiave che oscilla durante il ciclo di trasporto.
Dal legame lipidico al trasporto degli zuccheri
Queste differenze strutturali si traducono direttamente in funzione. I panorami di energia libera derivati da lunghe simulazioni mostrano che quando la cardiolipina è legata, il trasportatore può transitare con fluidità tra le conformazioni aperta verso l’esterno, occlusa e aperta verso l’interno necessarie per portare lo zucchero attraverso la membrana. Gli stati più stabili sono quelli rivolti verso l’esterno, in linea con la struttura sperimentale, ma il percorso verso le forme rivolte verso l’interno resta accessibile. Con il PG al comando, il quadro cambia: le sue code riempiono la cavità centrale, i contatti inter‑subunitari si disordinano e la proteina rimane intrappolata in una conformazione non produttiva in cui lo zucchero non può passare. La PE si comporta in modo simile al PG, favorendo l’accoppiamento delle subunità ma lasciandole più eterogenee e meno stabili rispetto allo stato legato alla cardiolipina.
Perché questo conta oltre un singolo trasportatore batterico
In termini pratici, questo lavoro mostra che i grassi di membrana non sono semplice lubrificante passivo ma parti meccaniche attive che possono attaccarsi alle proteine, sostenere pezzi in movimento o infilarsi in fessure critiche. Per VsSemiSWEET, la cardiolipina agisce come una spina sagomata su misura che sia blocca insieme le due metà sia guida i movimenti necessari al trasporto dello zucchero, mentre il PG tende a inceppare il meccanismo. Poiché molte proteine di membrana condividono siti lipidici interfaciali simili e spesso dipendono dalla cardiolipina o da lipidi correlati, questi risultati offrono uno schema più ampio su come la composizione lipidica possa modulare l’attività di trasportatori, canali e pompe in tutta la biologia.
Citazione: Zhang, Y., Zhao, W., Duan, M. et al. Diverse regulation of functional dimerization of a sugar transporter by different interfacial lipids. Nat Commun 17, 3062 (2026). https://doi.org/10.1038/s41467-026-69804-3
Parole chiave: lipidi di membrana, trasportatori di zuccheri, cardiolipina, dimerizzazione proteica, NMR in stato solido