Clear Sky Science · en
Diverse regulation of functional dimerization of a sugar transporter by different interfacial lipids
How Cell Fats Steer Tiny Sugar Gates
Our cells are packed with microscopic gatekeepers that move sugars and other nutrients across oily membranes. This study asks a deceptively simple question with big implications: can the surrounding fats themselves decide whether one such sugar gate forms the right shape and actually works? By watching a bacterial sugar transporter in a near‑natural membrane, the authors show that a few specific fat molecules at just the right spot can make the difference between a smooth-running gate and a jammed one.
The Sugar Gate at the Heart of the Story
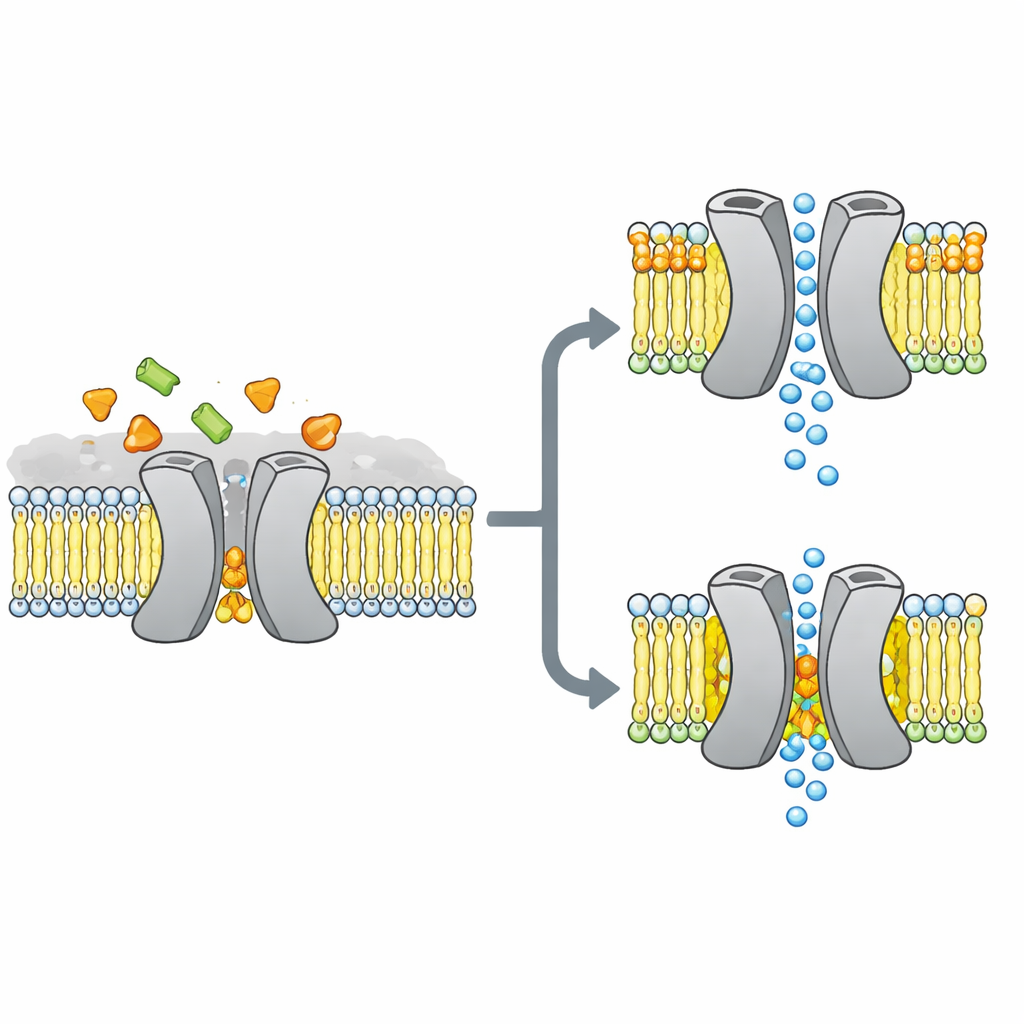
The researchers focused on VsSemiSWEET, a bacterial cousin of sugar transporters found in plants and animals. These proteins span the cell membrane and normally work as pairs, or dimers, to let sugar slip through from one side to the other. Earlier crystal structures had shown the overall shape of these pairs but curiously missed any bound fats, even though other methods suggested that certain lipids cling tightly to the transporter and might control how it behaves.
Finding the Hidden Fat Partners
To uncover which fats truly stick to this transporter in a lifelike setting, the team reconstituted VsSemiSWEET into membrane patches made from bacterial lipids and used solid‑state NMR, a powerful form of magnetic resonance that works on proteins inside intact membranes. They detected three co‑purifying lipid types: phosphatidylethanolamine (PE), phosphatidylglycerol (PG), and cardiolipin (CDL). All three nestle at the contact surface between the two protein partners, but cardiolipin binds especially tightly, surviving harsh detergent washes that strip away most other fats.
Seeing the Dimer in Its Native Habitat
Using a combination of carefully labeled samples and multidimensional NMR experiments, the authors built a high‑resolution 3D structure of the transporter dimer inside a lipid bilayer. They confirmed that the protein indeed forms a paired assembly with a cavity open toward the outside of the cell, suitable for sugar entry. When they mutated three positively charged amino acids at the dimer interface that contact lipids, the paired structure became fragile: in gels it fell apart more readily, and its NMR signals broadened and disappeared, indicating a flexible, unstable complex. Computer simulations echoed this picture, showing that without interfacial lipids, the two partners drift apart and lose many stabilizing contacts.
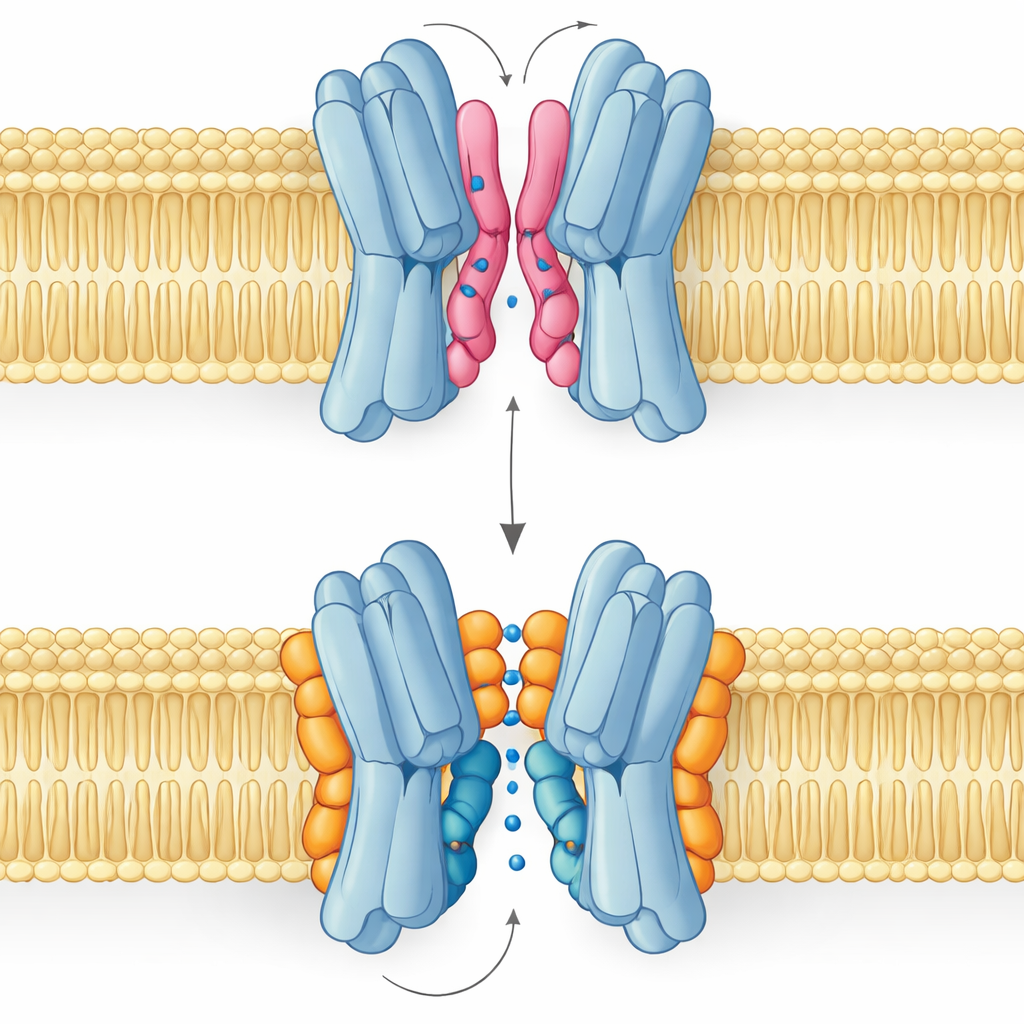
Good Lipid, Bad Lipid: A Tale of Two Modes of Binding
The most surprising result is that not all interface lipids help in the same way. Simulations and NMR measurements revealed that PG and cardiolipin bind at overlapping spots but adopt distinct poses. PG, with two tails and a single negative charge, can snake its tails deep into the narrow gap between the protein partners, prying them slightly apart and even plugging the central passage. Cardiolipin, by contrast, has a bulkier head carrying two negative charges and four tails. Its head grips a cluster of basic residues like a clamp, while its larger tail bundle stays on the outside surface, avoiding the central cavity and instead bracing one of the key helices that rocks during the transport cycle.
From Lipid Binding to Sugar Transport
These structural differences translate directly into function. Free‑energy landscapes from long simulations show that when cardiolipin is bound, the transporter can smoothly visit the outward‑open, occluded, and inward‑open shapes needed to carry sugar across the membrane. The most stable states are outward‑facing, in line with the experimental structure, but the pathway to inward‑facing forms remains accessible. With PG in control, the picture changes: its tails fill the central cavity, inter‑subunit contacts become disordered, and the protein is trapped in a non‑productive conformation where sugar cannot pass. PE behaves similarly to PG, promoting pairing of the subunits but leaving them more heterogeneous and less stable than in the cardiolipin‑bound state.
Why This Matters Beyond One Bacterial Transporter
In everyday terms, this work shows that membrane fats are not just passive grease but active mechanical parts that can latch onto proteins, brace moving pieces, or wedge into critical gaps. For VsSemiSWEET, cardiolipin acts like a custom‑shaped shim that both locks the two halves together and guides the motions needed for sugar transport, while PG tends to gum up the works. Because many membrane proteins share similar interfacial lipid sites and often rely on cardiolipin or related lipids, these findings offer a broader blueprint for how changing lipid composition could tune the activity of transporters, channels, and pumps across biology.
Citation: Zhang, Y., Zhao, W., Duan, M. et al. Diverse regulation of functional dimerization of a sugar transporter by different interfacial lipids. Nat Commun 17, 3062 (2026). https://doi.org/10.1038/s41467-026-69804-3
Keywords: membrane lipids, sugar transporters, cardiolipin, protein dimerization, solid-state NMR