Clear Sky Science · es
Regulación diversa de la dimerización funcional de un transportador de azúcares por diferentes lípidos interfaciales
Cómo las grasas celulares dirigen pequeñas compuertas de azúcar
Nuestras células están llenas de guardias microscópicos que mueven azúcares y otros nutrientes a través de membranas oleosas. Este estudio plantea una pregunta aparentemente simple con grandes implicaciones: ¿pueden las propias grasas circundantes decidir si una de esas compuertas de azúcar adopta la forma correcta y funciona realmente? Al observar un transportador bacteriano de azúcares en una membrana casi natural, los autores muestran que unas pocas moléculas de grasa específicas en el lugar preciso pueden marcar la diferencia entre una compuerta que funciona con fluidez y otra que se atasca.
La compuerta de azúcar en el centro de la historia
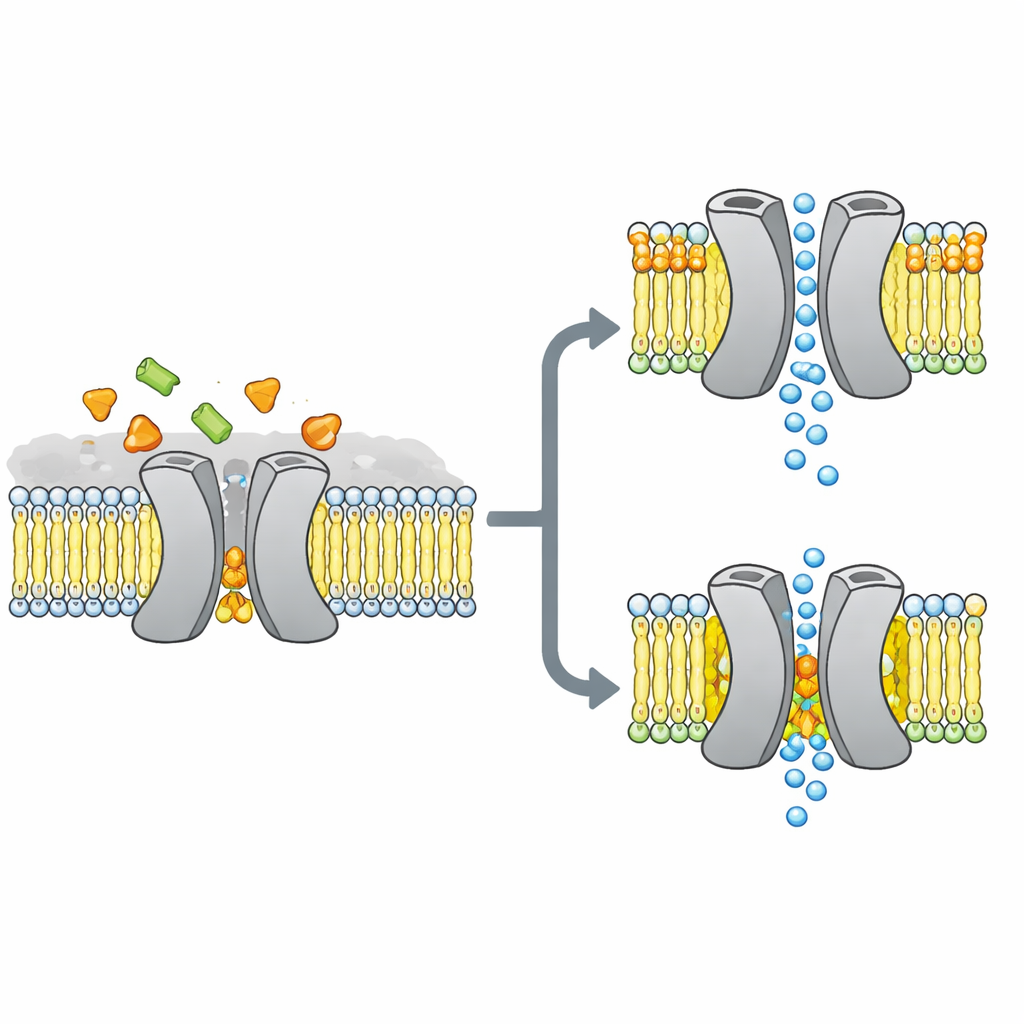
Los investigadores se centraron en VsSemiSWEET, un pariente bacteriano de los transportadores de azúcares que se encuentran en plantas y animales. Estas proteínas atraviesan la membrana celular y normalmente funcionan en parejas, o dímeros, para permitir que el azúcar pase de un lado al otro. Estructuras cristalográficas previas habían mostrado la forma general de estos pares, pero curiosamente no detectaron lípidos unidos, a pesar de que otros métodos sugerían que ciertos lípidos se adhieren fuertemente al transportador y podrían controlar su comportamiento.
Encontrar a los socios lipídicos ocultos
Para descubrir qué grasas se adhieren realmente a este transportador en un entorno vivo, el equipo reconstituyó VsSemiSWEET en parches de membrana hechos con lípidos bacterianos y empleó RMN en estado sólido, una potente forma de resonancia magnética que funciona sobre proteínas dentro de membranas intactas. Detectaron tres tipos de lípidos que co‑purificaban: fosfatidiletanolamina (PE), fosfatidilglicerol (PG) y cardiolipina (CDL). Los tres se alojan en la superficie de contacto entre las dos subunidades proteicas, pero la cardiolipina se une de forma especialmente fuerte, resistiendo lavados con detergente agresivos que eliminan la mayoría de las otras grasas.
Ver el dímero en su hábitat nativo
Usando una combinación de muestras cuidadosamente etiquetadas y experimentos multidimensionales de RMN, los autores construyeron una estructura 3D de alta resolución del dímero del transportador dentro de una bicapa lipídica. Confirmaron que la proteína efectivamente forma un ensamblaje pareado con una cavidad abierta hacia el exterior celular, adecuada para la entrada de azúcar. Cuando mutaron tres aminoácidos cargados positivamente en la interfaz del dímero que contactan lípidos, la estructura pareada se volvió frágil: en geles se disociaba con más facilidad y sus señales de RMN se ensanchaban y desaparecían, lo que indica un complejo flexible e inestable. Simulaciones por ordenador replicaron este panorama, mostrando que sin lípidos interfaciales, las dos subunidades se separan y pierden muchos contactos estabilizadores.
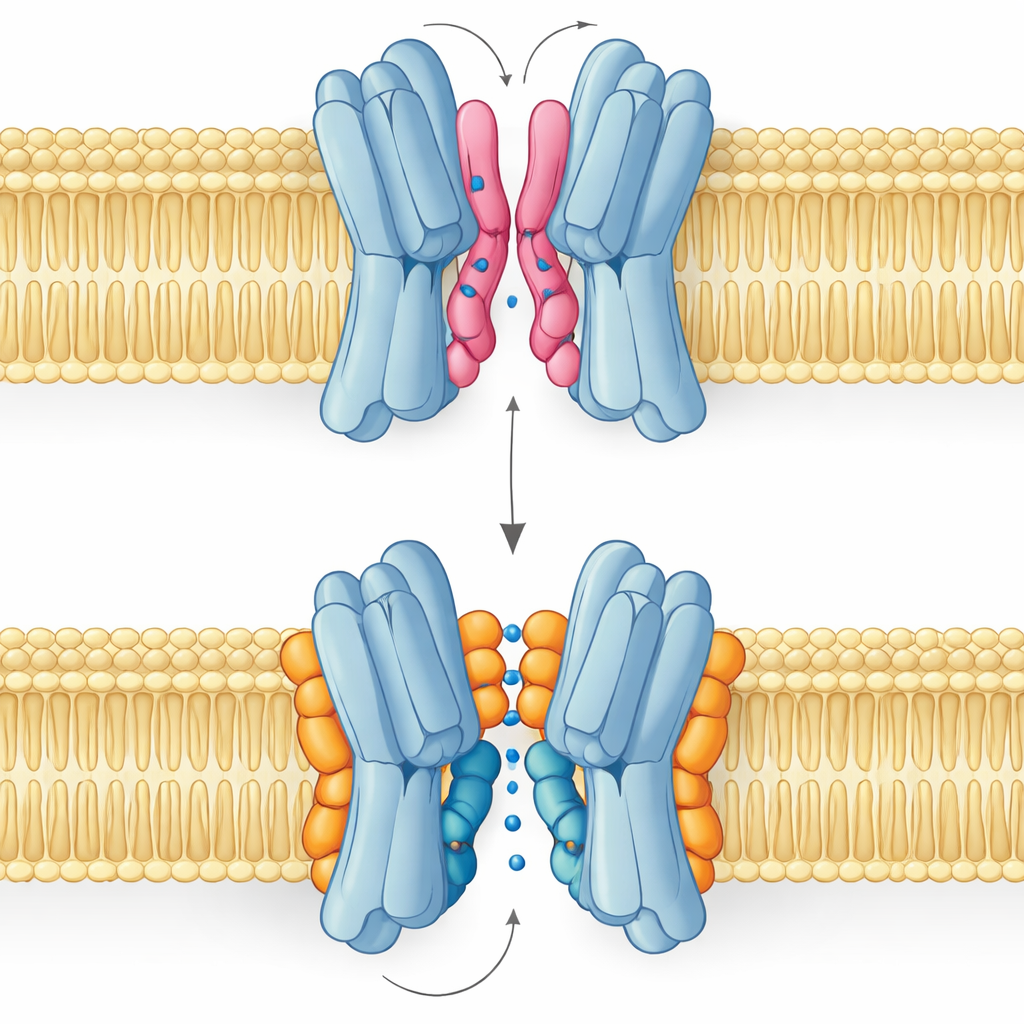
Buen lípido, mal lípido: la historia de dos modos de unión
El resultado más sorprendente es que no todos los lípidos de la interfaz ayudan de la misma manera. Simulaciones y mediciones por RMN revelaron que PG y cardiolipina se unen en puntos solapados pero adoptan posturas distintas. El PG, con dos colas y una sola carga negativa, puede introducir sus colas profundamente en la estrecha hendidura entre las subunidades, separándolas ligeramente e incluso taponando el pasaje central. La cardiolipina, por el contrario, tiene una cabeza más voluminosa con dos cargas negativas y cuatro colas. Su cabeza sujeta un racimo de residuos básicos como una abrazadera, mientras que su mayor haz de colas permanece en la superficie exterior, evitando la cavidad central y en su lugar apuntalando una de las hélices clave que se balancea durante el ciclo de transporte.
De la unión lipídica al transporte de azúcares
Estas diferencias estructurales se traducen directamente en la función. Los paisajes de energía libre obtenidos de simulaciones largas muestran que cuando la cardiolipina está unida, el transportador puede visitar con fluidez las conformaciones abierto hacia afuera, ocluido y abierto hacia adentro necesarias para llevar el azúcar a través de la membrana. Los estados más estables son los orientados hacia el exterior, en consonancia con la estructura experimental, pero la vía hacia las formas orientadas hacia el interior sigue siendo accesible. Con PG presente, el panorama cambia: sus colas llenan la cavidad central, los contactos entre subunidades se desordenan y la proteína queda atrapada en una conformación no productiva donde el azúcar no puede pasar. La PE se comporta de manera similar al PG, promoviendo el apareamiento de las subunidades pero dejándolas más heterogéneas y menos estables que en el estado unido a cardiolipina.
Por qué esto importa más allá de un transportador bacteriano
En términos cotidianos, este trabajo muestra que las grasas de membrana no son solo lubricante pasivo sino piezas mecánicas activas que pueden engancharse a proteínas, apuntalar partes móviles o clavarse en huecos críticos. Para VsSemiSWEET, la cardiolipina actúa como una cuña de forma personalizada que tanto bloquea las dos mitades como guía los movimientos necesarios para el transporte de azúcares, mientras que el PG tiende a entorpecer el funcionamiento. Dado que muchas proteínas de membrana comparten sitios lipídicos interfaciales similares y a menudo dependen de cardiolipina o lípidos relacionados, estos hallazgos ofrecen un esquema más amplio de cómo cambiar la composición lipídica podría ajustar la actividad de transportadores, canales y bombas en la biología.»}
Cita: Zhang, Y., Zhao, W., Duan, M. et al. Diverse regulation of functional dimerization of a sugar transporter by different interfacial lipids. Nat Commun 17, 3062 (2026). https://doi.org/10.1038/s41467-026-69804-3
Palabras clave: lípidos de membrana, transportadores de azúcares, cardiolipina, dimerización de proteínas, RMN en estado sólido