Clear Sky Science · fr
Régulation diverse de la dimérisation fonctionnelle d’un transporteur de sucre par différentes lipides interfaciaux
Comment les graisses cellulaires orientent de minuscules portails à sucre
Nos cellules regorgent de gardiens microscopiques qui déplacent les sucres et autres nutriments à travers des membranes huileuses. Cette étude pose une question apparemment simple mais lourde de conséquences : les graisses environnantes peuvent‑elles, à elles seules, décider si l’un de ces portails à sucre prend la bonne forme et fonctionne réellement ? En observant un transporteur de sucre bactérien dans une membrane proche du naturel, les auteurs montrent que quelques molécules lipidiques spécifiques, placées au bon endroit, peuvent faire la différence entre un portail qui fonctionne sans accroc et un portail bloqué.
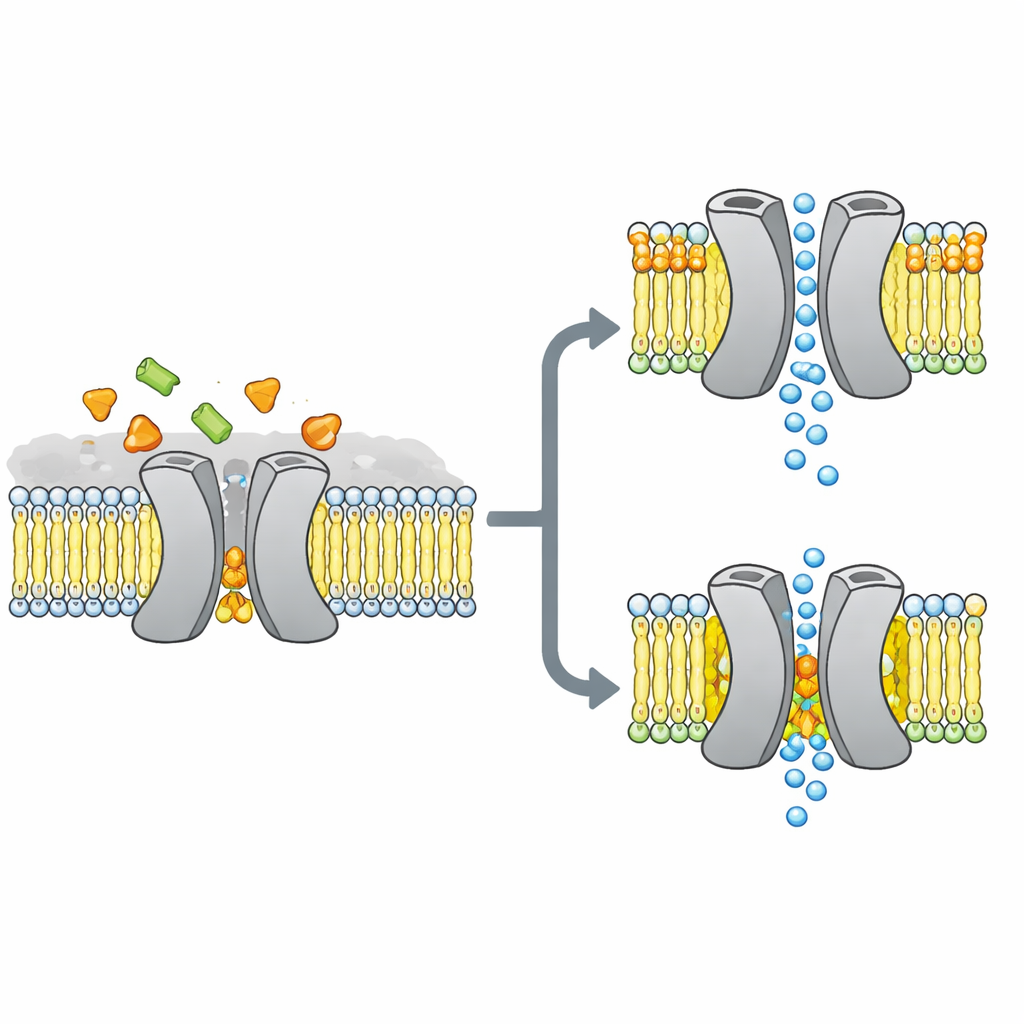
Le portail à sucre au cœur de l’histoire
Les chercheurs se sont concentrés sur VsSemiSWEET, un cousin bactérien des transporteurs de sucre présents chez les plantes et les animaux. Ces protéines traversent la membrane cellulaire et fonctionnent normalement par paires, ou dimères, pour laisser le sucre glisser d’un côté à l’autre. Des structures cristallographiques antérieures avaient montré la forme globale de ces paires mais, curieusement, n’avaient pas révélé de lipides liés, alors que d’autres méthodes suggéraient que certains lipides s’attachent fermement au transporteur et pourraient contrôler son comportement.
Trouver les partenaires lipidiques cachés
Pour découvrir quels lipides restent réellement associés à ce transporteur dans un cadre proche du vivant, l’équipe a reconstitué VsSemiSWEET dans des patchs membranaires faits de lipides bactériens et a utilisé la RMN en état solide, une forme puissante de résonance magnétique applicable aux protéines dans des membranes intactes. Ils ont détecté trois types de lipides co‑purifiants : la phosphatidyléthanolamine (PE), la phosphatidylglycérol (PG) et la cardiolipine (CDL). Les trois se logent à la surface de contact entre les deux partenaires protéiques, mais la cardiolipine se lie particulièrement fortement, résistant à des lavages au détergent sévères qui éliminent la plupart des autres lipides.
Voir le dimère dans son habitat natif
En combinant des échantillons soigneusement marqués et des expériences RMN multidimensionnelles, les auteurs ont construit une structure 3D haute résolution du dimère du transporteur au sein d’une bicouche lipidique. Ils ont confirmé que la protéine forme bien une assemblée par paire avec une cavité ouverte vers l’extérieur de la cellule, adaptée à l’entrée du sucre. Lorsque trois acides aminés chargés positivement à l’interface du dimère en contact avec les lipides ont été mutés, la structure dimérique est devenue fragile : en gel elle se dissociait plus facilement, et ses signaux RMN s’élargissaient puis disparaissaient, indiquant un complexe flexible et instable. Des simulations informatiques vont dans le même sens, montrant que sans lipides interfaciaux, les deux partenaires dérivent l’un par rapport à l’autre et perdent de nombreux contacts stabilisateurs.
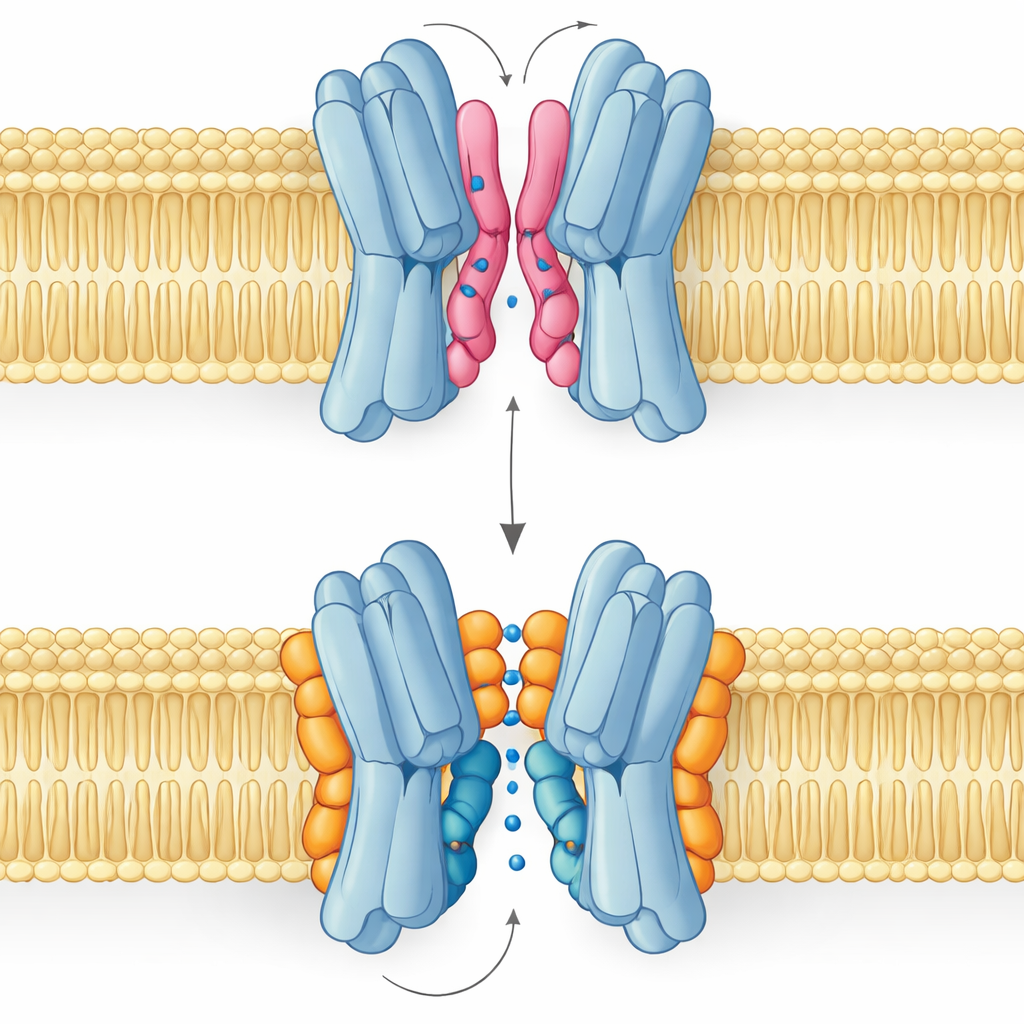
Bon lipide, mauvais lipide : l’histoire de deux modes de liaison
Le résultat le plus surprenant est que tous les lipides d’interface n’aident pas de la même manière. Les simulations et les mesures RMN ont révélé que la PG et la cardiolipine se lient à des emplacements chevauchants mais adoptent des poses distinctes. La PG, avec deux queues et une charge négative simple, peut enfoncer ses queues profondément dans l’étroite fente entre les partenaires protéiques, les écartant légèrement et même boucher le passage central. La cardiolipine, en revanche, possède une tête plus volumineuse portant deux charges négatives et quatre queues. Sa tête saisit un groupe de résidus basiques comme une pince, tandis que son faisceau de queues plus large reste sur la surface externe, évitant la cavité centrale et venant plutôt consolider l’une des hélices clés qui bascule pendant le cycle de transport.
De la liaison lipidique au transport du sucre
Ces différences structurelles se traduisent directement en fonction. Les paysages d’énergie libre issus de longues simulations montrent que lorsque la cardiolipine est liée, le transporteur peut parcourir sans encombre les états ouvert vers l’extérieur, occlus et ouvert vers l’intérieur nécessaires pour acheminer le sucre à travers la membrane. Les états les plus stables sont orientés vers l’extérieur, en accord avec la structure expérimentale, mais la voie vers les conformations orientées vers l’intérieur reste accessible. Sous l’influence de la PG, le tableau change : ses queues remplissent la cavité centrale, les contacts inter‑sous‑unités deviennent désordonnés et la protéine reste piégée dans une conformation non productive où le sucre ne peut pas passer. Le PE se comporte de manière similaire à la PG, favorisant la mise en paire des sous‑unités mais les laissant plus hétérogènes et moins stables que dans l’état lié à la cardiolipine.
Pourquoi cela importe au‑delà d’un transporteur bactérien
En termes simples, ce travail montre que les graisses membranaires ne sont pas de la simple graisse passive mais des pièces mécaniques actives qui peuvent s’accrocher aux protéines, caler des éléments mobiles ou se glisser dans des interstices critiques. Pour VsSemiSWEET, la cardiolipine agit comme une cale sur mesure qui verrouille les deux moitiés ensemble et guide les mouvements nécessaires au transport du sucre, tandis que la PG tend à enrayer le fonctionnement. Parce que de nombreuses protéines membranaires partagent des sites lipidiques interfaciaux similaires et reposent souvent sur la cardiolipine ou des lipides apparentés, ces résultats proposent un cadre plus général pour comprendre comment la composition lipidique peut moduler l’activité de transporteurs, canaux et pompes dans la biologie.
Citation: Zhang, Y., Zhao, W., Duan, M. et al. Diverse regulation of functional dimerization of a sugar transporter by different interfacial lipids. Nat Commun 17, 3062 (2026). https://doi.org/10.1038/s41467-026-69804-3
Mots-clés: lipides membranaires, transporteurs de sucre, cardiolipine, dimérisation des protéines, RMN en état solide