Clear Sky Science · ru
Разнообразная регуляция функциональной димеризации переносчика сахара разными интерфейсными липидами
Как клеточные жиры управляют крошечными сахарными воротами
В наших клетках полно крошечных стражей, которые переносят сахара и другие питательные вещества через масляные мембраны. В этом исследовании ставится на первый взгляд простой, но важный вопрос: могут ли сами окружающие жиры решить, образует ли такой переносчик сахара правильную форму и действительно ли он работает? Наблюдая за бактериальным переносчиком сахара в максимально естественной мембране, авторы показывают, что несколько конкретных молекул жира в нужном месте могут решить, будет ли ворота работать плавно или заедать.
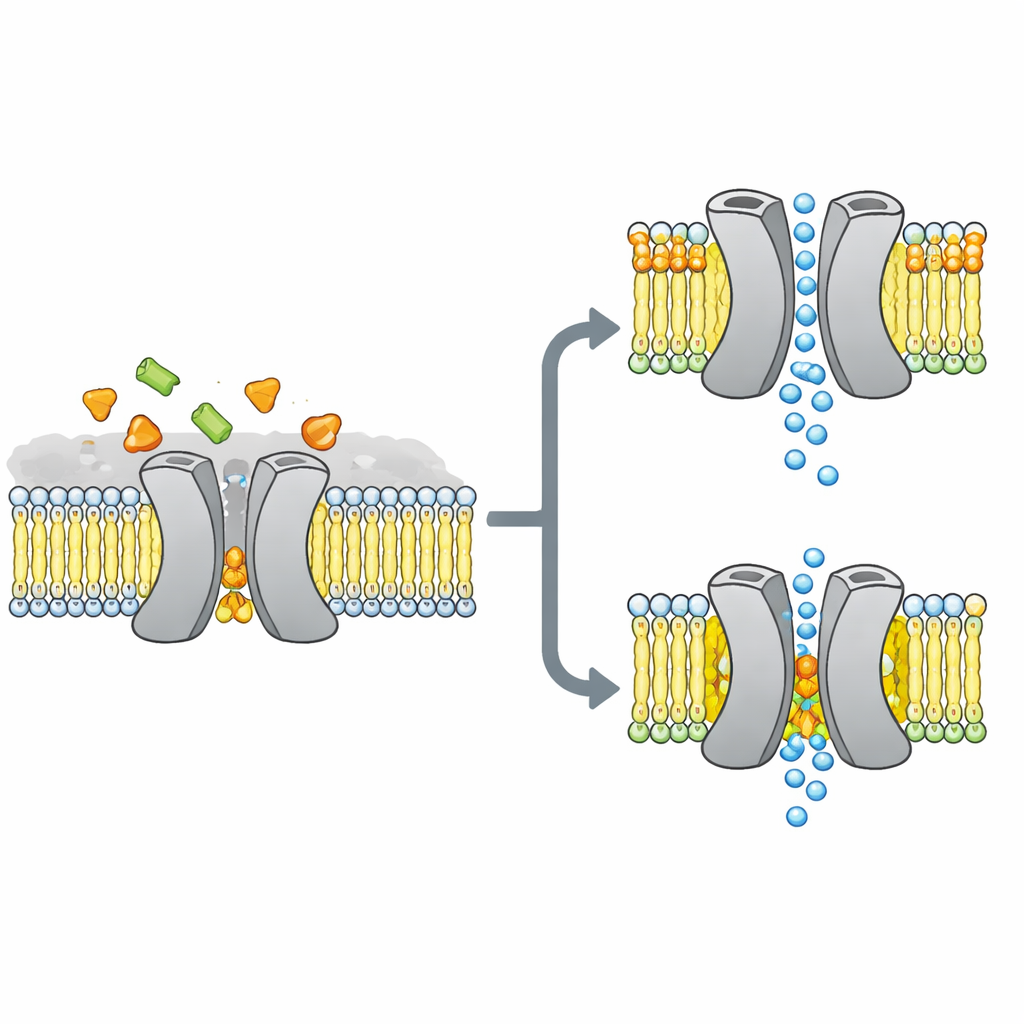
Сахарные ворота в центре истории
Исследователи сосредоточились на VsSemiSWEET, бактериальном родственнике переносчиков сахара, встречающихся у растений и животных. Эти белки пронизывают клеточную мембрану и обычно работают парами, или димерами, позволяя сахару проскакивать с одной стороны на другую. Ранее кристаллические структуры показали общую форму этих пар, но, любопытно, не продемонстрировали связанных липидов, хотя другие методы указывали, что определённые липиды прочно прилипают к переносчику и могут контролировать его поведение.
Поиск скрытых липидных партнёров
Чтобы выяснить, какие липиды действительно остаются с этим переносчиком в приближённых к живым условиях, команда реконсититуировала VsSemiSWEET в мембранные пласты из бактериальных липидов и использовала твёрдотельный ЯМР — мощный метод магнитного резонанса, работающий с белками в целых мембранах. Они обнаружили три типа липидов, сосуществующих с белком: фосфатидилэтаноламин (PE), фосфатидилглицерин (PG) и кардиолипин (CDL). Все три располагаются у контактной поверхности между двумя белковыми партнёрами, но кардиолипин связывается особенно прочно, выдерживая жёсткие детергентные промывки, которые смывают большинство других липидов.
Видеть димер в его естественной среде
Используя сочетание тщательно меченых образцов и многомерных ЯМР‑экспериментов, авторы построили высокоразрешающую 3D‑структуру димера переносчика внутри липидного бислоя. Они подтвердили, что белок действительно образует парную сборку с полостью, открытой к внешней стороне клетки, пригодной для входа сахара. При замене трёх положительно заряженных аминокислот на интерфейсе димера, контактирующих с липидами, парная структура становилась хрупкой: в гелях она легче распадалась, а её сигналы в ЯМР расширялись и исчезали, что указывает на гибкий, нестабильный комплекс. Компьютерные симуляции подтвердили эту картину: без интерфейсных липидов два партнёра расходятся и теряют многие стабилизирующие контакты.
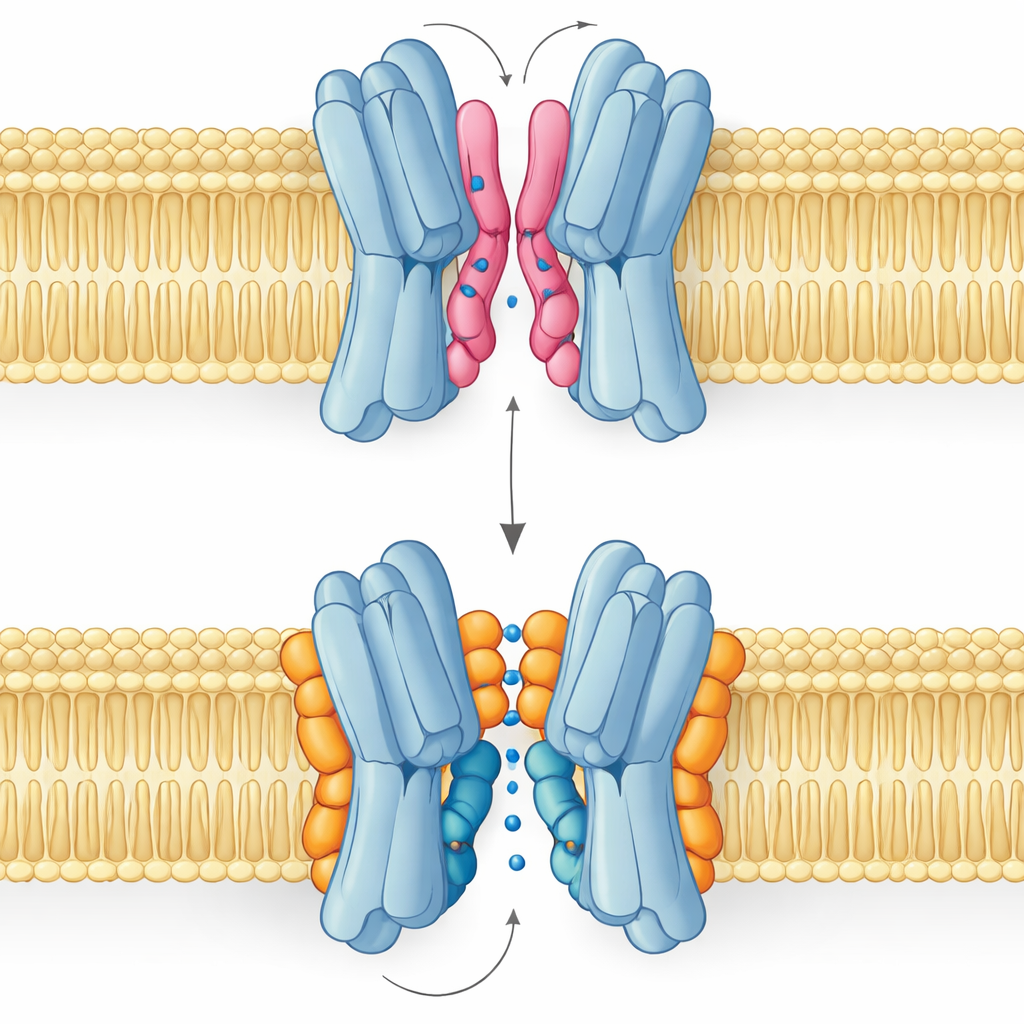
Хороший липид, плохой липид: история двух режимов связывания
Самый неожиданный результат состоит в том, что не все липиды интерфейса помогают одинаково. Симуляции и ЯМР‑измерения показали, что PG и кардиолипин связываются в перекрывающихся участках, но занимают разные позы. PG, с двумя хвостами и одним отрицательным зарядом, может засовывать свои хвосты глубоко в узкий зазор между белковыми партнёрами, слегка раздвигая их и даже закупоривая центральный проход. Кардиолипин, напротив, имеет более массивную головку с двумя отрицательными зарядами и четыре хвоста. Его головка захватывает кластер основательных остатков как зажим, в то время как большая связка хвостов остаётся на внешней поверхности, избегая центральной полости и подпиная одну из ключевых спиралей, которая качается во время транспортного цикла.
От связывания липидов к переносу сахара
Эти структурные различия прямо влияют на функцию. Ландшафты свободной энергии из долгих симуляций показывают, что при связывании кардиолипина переносчик может плавно переходить между внешне открытыми, окклюзированными и внутренне открытыми формами, необходимыми для переноса сахара через мембрану. Наиболее стабильными состояниями являются внешне ориентированные, что согласуется с экспериментальной структурой, но путь к внутренним формам остаётся доступным. Под контролем PG картина меняется: его хвосты заполняют центральную полость, межсубъединичные контакты разупорядочиваются, и белок оказывается в непродуктивной конформации, где сахар не может пройти. PE ведёт себя схоже с PG, способствуя парированию субъединиц, но делая их более гетерогенными и менее стабильными, чем в состоянии, связанном с кардиолипином.
Почему это важно не только для одного бактериального переносчика
Проще говоря, эта работа показывает, что мембранные жиры — не просто пассивная смазка, а активные механические части, которые могут цепляться за белки, подпирать подвижные элементы или втыкаться в критические зазоры. Для VsSemiSWEET кардиолипин действует как специально сформованная прокладка, которая одновременно фиксирует две половины и направляет движения, необходимые для переноса сахара, тогда как PG склонен заклинить механизм. Поскольку многие мембранные белки имеют схожие интерфейсные липидные участки и часто зависят от кардиолипина или родственных липидов, эти результаты дают более широкую основу для понимания того, как изменение состава липидов может настраивать активность переносчиков, каналов и насосов в биологии.
Цитирование: Zhang, Y., Zhao, W., Duan, M. et al. Diverse regulation of functional dimerization of a sugar transporter by different interfacial lipids. Nat Commun 17, 3062 (2026). https://doi.org/10.1038/s41467-026-69804-3
Ключевые слова: мембранные липиды, переносчики сахаров, кардиолипин, димеризация белков, твёрдотельный ЯМР