Clear Sky Science · de
Verschiedene Regulation der funktionellen Dimerisierung eines Zuckertransporters durch verschiedene interfaciale Lipide
Wie Zellfette winzige Zuckertore steuern
Unsere Zellen sind voller mikroskopischer Wächter, die Zucker und andere Nährstoffe durch ölige Membranen transportieren. Diese Studie stellt eine auf den ersten Blick einfache, aber weitreichende Frage: Können die umgebenden Fette selbst darüber entscheiden, ob ein solcher Zuckerkorridor die richtige Form annimmt und tatsächlich funktioniert? Indem die Autoren einen bakteriellen Zuckertransporter in einer nahezu natürlichen Membran beobachteten, zeigen sie, dass wenige spezifische Fettmoleküle an genau der richtigen Stelle den Unterschied zwischen einem reibungslos arbeitenden Tor und einem blockierten ausmachen können.
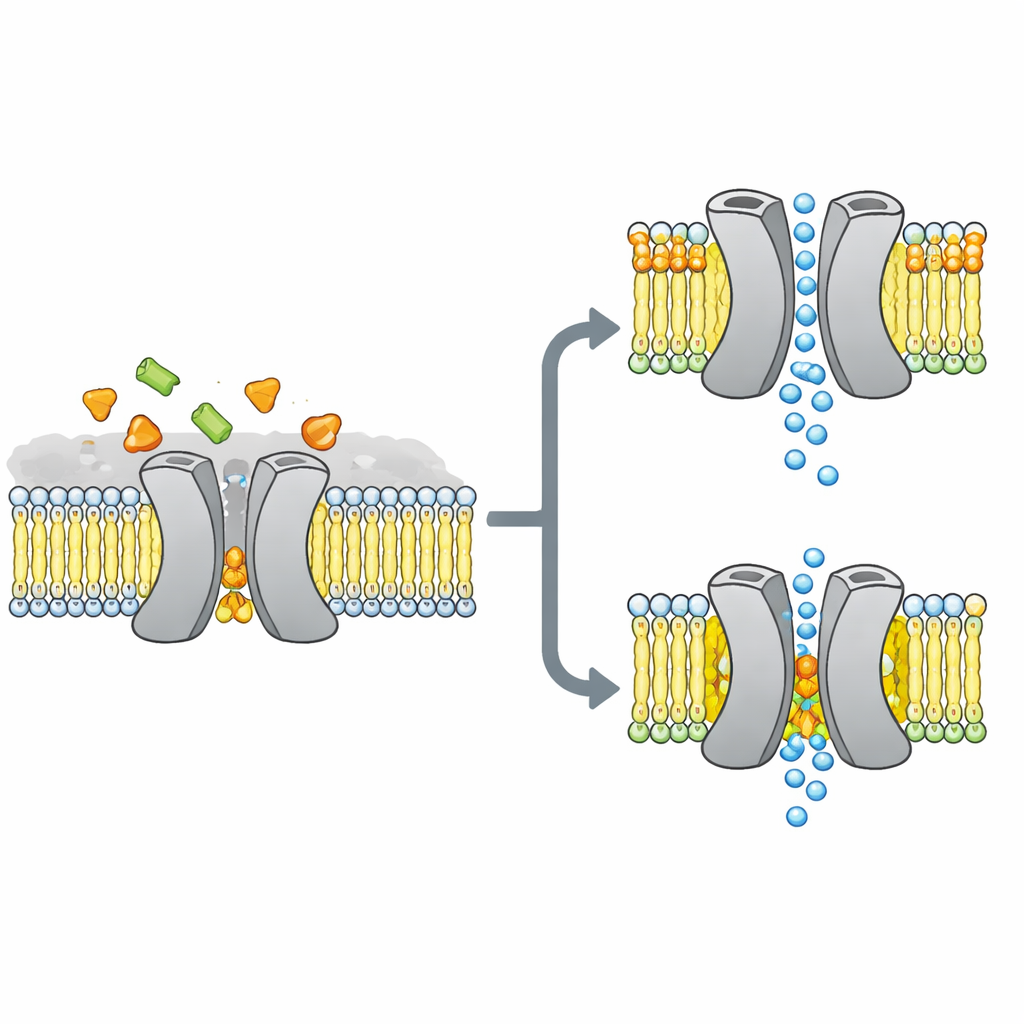
Das Zuckertor im Mittelpunkt der Geschichte
Die Forscher konzentrierten sich auf VsSemiSWEET, einen bakteriellen Verwandten von Zuckertransportern, die auch in Pflanzen und Tieren vorkommen. Diese Proteine durchspannen die Zellmembran und arbeiten normalerweise als Paare, sogenannte Dimere, um Zucker von einer Seite zur anderen zu lassen. Frühere Kristallstrukturen zeigten die Gesamtform dieser Paare, vermissten aber merkwürdigerweise gebundene Lipide, obwohl andere Methoden nahelegten, dass bestimmte Lipide fest an den Transporter binden und sein Verhalten steuern könnten.
Die verborgenen Fettpartner finden
Um aufzudecken, welche Lipide in einer lebensnahen Umgebung tatsächlich an diesen Transporter binden, setzten die Forscher VsSemiSWEET in Membranfragmente aus bakteriellen Lipiden zurück und nutzten Festkörper‑NMR, eine leistungsfähige Form der Magnetresonanz, die an Proteinen in intakten Membranen arbeitet. Sie entdeckten drei mitreinigende Lipidtypen: Phosphatidylethanolamin (PE), Phosphatidylglycerol (PG) und Kardiolipin (CDL). Alle drei liegen an der Kontaktfläche zwischen den beiden Proteinpartnern, aber Kardiolipin bindet besonders stark und übersteht sogar harte Detergent‑Wäschen, die die meisten anderen Lipide entfernen.
Das Dimer in seiner natürlichen Umgebung sehen
Mithilfe einer Kombination aus sorgfältig markierten Proben und multidimensionalen NMR‑Experimenten bauten die Autoren eine hochaufgelöste 3D‑Struktur des Transporter‑Dimers innerhalb einer Lipiddoppelschicht auf. Sie bestätigten, dass das Protein tatsächlich eine gepaarte Anordnung bildet mit einer nach außen offenen Taschen, die für das Eindringen von Zucker geeignet ist. Wenn sie drei positiv geladene Aminosäuren an der Dimeroberfläche, die Lipide kontaktieren, mutierten, wurde die Paarstruktur brüchig: in Gelen fiel sie leichter auseinander und ihre NMR‑Signale wurden breiter und verschwanden, ein Hinweis auf einen flexiblen, instabilen Komplex. Computersimulationen bestätigten dieses Bild und zeigten, dass ohne interfaciale Lipide die beiden Partner auseinanderdriften und viele stabilisierende Kontakte verlieren.
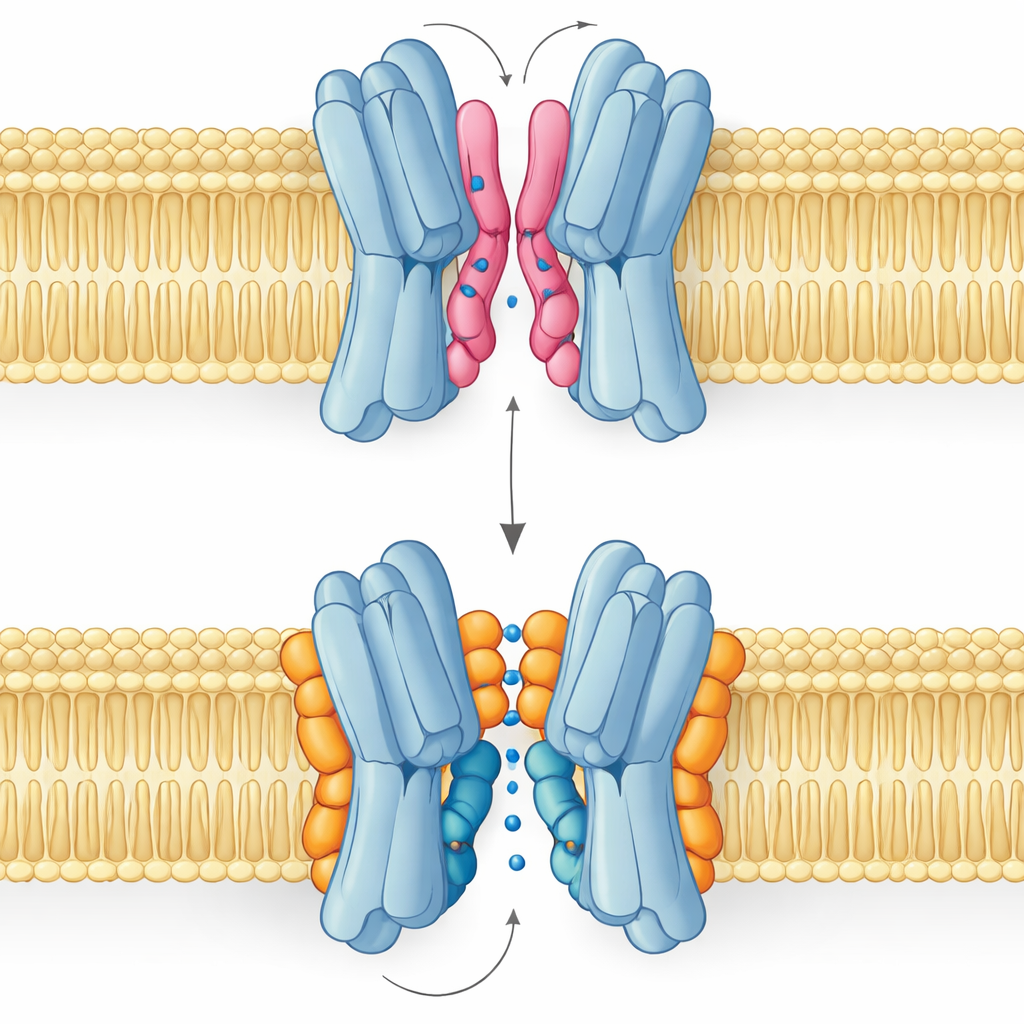
Gutes Lipid, schlechtes Lipid: Zwei Bindungsarten
Das überraschendste Ergebnis ist, dass nicht alle Interfacialipide auf die gleiche Weise helfen. Simulationen und NMR‑Messungen zeigten, dass PG und Kardiolipin an sich überlappenden Stellen binden, dabei aber unterschiedliche Haltungen einnehmen. PG, mit zwei Schwänzen und einer negativen Ladung, kann seine Schwänze tief in die enge Lücke zwischen den Proteinpartnern schieben, sie leicht auseinandertreiben und sogar den zentralen Durchgang verstopfen. Kardiolipin dagegen hat einen voluminöseren Kopf mit zwei negativen Ladungen und vier Schwänzen. Sein Kopf greift eine Gruppe basischer Reste wie eine Klammer, während sein größeres Schwanzbündel an der Außenseite bleibt, die zentrale Kavität meidet und stattdessen eine der Schlüsselhelices abstützt, die sich während des Transportzyklus bewegt.
Von Lipidbindung zum Zuckerverkehr
Diese strukturellen Unterschiede wirken sich direkt auf die Funktion aus. Freie‑Energie‑Landkarten aus langen Simulationen zeigen, dass der Transporter mit gebundenem Kardiolipin reibungslos die nach außen offene, okkludierte und nach innen offene Konformationen durchlaufen kann, die zum Transport von Zucker nötig sind. Die stabilsten Zustände sind nach außen gerichtet, im Einklang mit der experimentellen Struktur, aber der Weg zu nach innen gerichteten Formen bleibt zugänglich. Unter dem Einfluss von PG ändert sich das Bild: seine Schwänze füllen die zentrale Kavität, die inter‑Subunit‑Kontakte werden ungeordnet und das Protein sitzt in einer nicht‑produktiven Konformation fest, durch die Zucker nicht passieren kann. PE verhält sich ähnlich wie PG: es fördert zwar das Zusammenfinden der Untereinheiten, lässt sie aber heterogener und weniger stabil zurück als im Kardiolipin‑gebundenen Zustand.
Warum das über einen einzelnen bakteriellen Transporter hinaus wichtig ist
Alltäglich ausgedrückt zeigt diese Arbeit, dass Membranlipide nicht nur passive Schmierstoffe sind, sondern aktive mechanische Bauteile, die an Proteine andocken, bewegliche Teile abstützen oder sich in kritische Spalte schieben können. Für VsSemiSWEET wirkt Kardiolipin wie eine maßgeschneiderte Unterlage, die beide Hälften zusammen verriegelt und gleichzeitig die Bewegungen lenkt, die für den Zuckertransport nötig sind, während PG dazu neigt, die Funktion zu blockieren. Da viele Membranproteine ähnliche interfaciale Lipidstellen haben und oft auf Kardiolipin oder verwandte Lipide angewiesen sind, liefern diese Ergebnisse eine breitere Blaupause dafür, wie Veränderungen der Lipidzusammensetzung die Aktivität von Transportern, Kanälen und Pumpen in der Biologie feinabstimmen könnten.
Zitation: Zhang, Y., Zhao, W., Duan, M. et al. Diverse regulation of functional dimerization of a sugar transporter by different interfacial lipids. Nat Commun 17, 3062 (2026). https://doi.org/10.1038/s41467-026-69804-3
Schlüsselwörter: Membranlipide, Zuckertransporter, Kardiolipin, Proteindimerisierung, Festkörper‑NMR