Clear Sky Science · pl
Różnorodna regulacja funkcjonalnej dymeryzacji transportera cukrów przez różne lipidy interfejsu
Jak tłuszcze komórkowe kierują mikroskopijnymi wrótami cukrowymi
Nasze komórki są wypełnione mikroskopijnymi strażnikami, które przenoszą cukry i inne składniki odżywcze przez oleiste błony. To badanie stawia na pozór proste pytanie o dalekosiężnych konsekwencjach: czy otaczające tłuszcze same mogą zadecydować, czy taki kanał cukrowy przyjmie właściwy kształt i będzie działał? Obserwując bakteryjny transporter cukrów w warunkach zbliżonych do naturalnych, autorzy pokazują, że kilka konkretnych cząsteczek tłuszczu umieszczonych w odpowiednim miejscu może przesądzić, czy wrótka działają płynnie, czy są zablokowane.
Wrota cukrowe w centrum opowieści
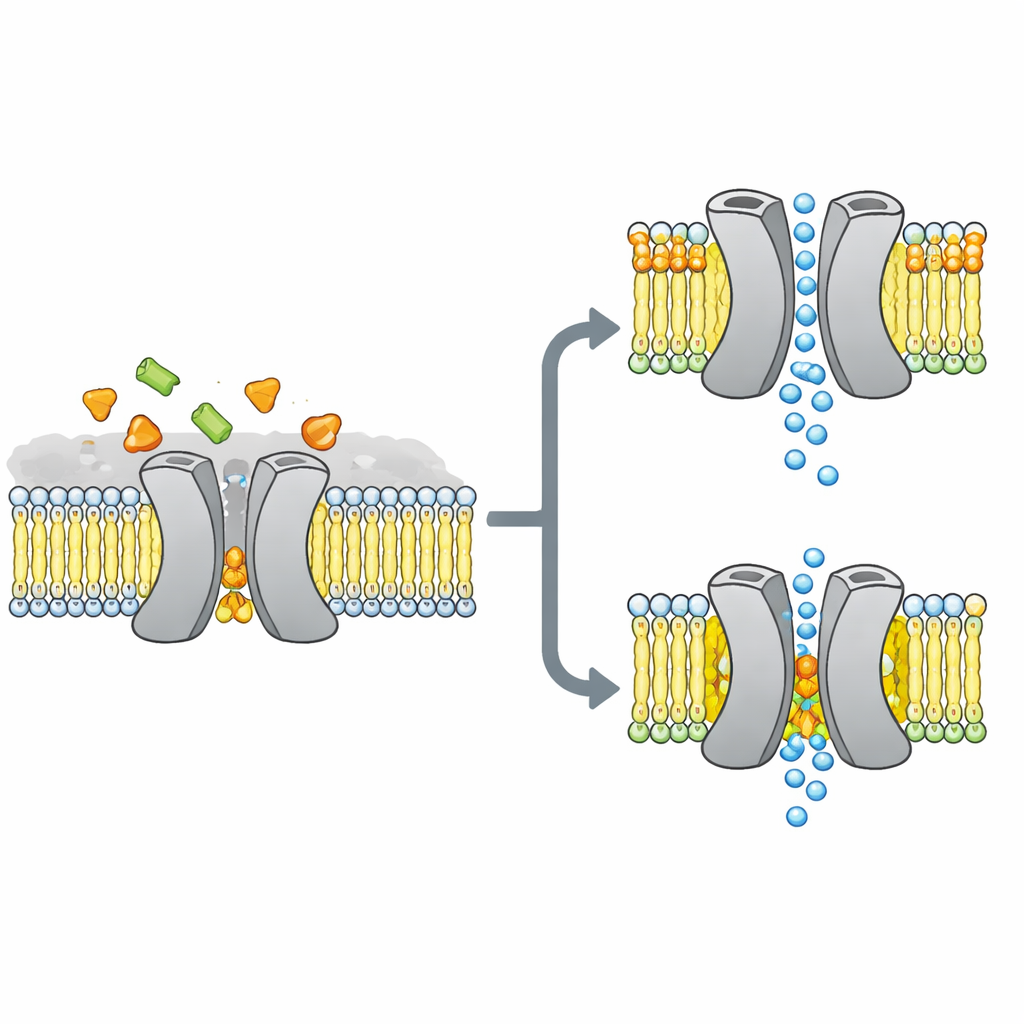
Badacze skupili się na VsSemiSWEET, bakteryjnym kuzynie transporterów cukrów występujących u roślin i zwierząt. Te białka przecinają błonę komórkową i zwykle działają w parach, czyli dimerach, pozwalając cukrom przemieszczać się z jednej strony na drugą. Wcześniejsze struktury krystaliczne ujawniały ogólny kształt tych par, ale ciekawie pomijały związane lipidy, mimo że inne metody sugerowały, iż pewne lipidy mocno przywierają do transportera i mogą kontrolować jego zachowanie.
Odkrywanie ukrytych partnerów tłuszczowych
Aby odsłonić, które tłuszcze rzeczywiście przylegają do tego transportera w warunkach bliskich żywym, zespół rekalibrował VsSemiSWEET do fragmentów błony zrobionych z bakteryjnych lipidów i zastosował NMR ciała stałego — potężną formę rezonansu magnetycznego działającą na białka wewnątrz nienaruszonych błon. Wykryli trzy typy lipidów ko‑purujących się z białkiem: fosfatydyloetanolaminę (PE), fosfatydyloglicerol (PG) oraz kardiolipinę (CDL). Wszystkie trzy gnieżdżą się na powierzchni styku między dwoma partnerami białkowymi, ale kardiolipina wiąże się szczególnie mocno, przetrzymując ostre prania detergentem, które zmywają większość innych tłuszczów.
Widząc dimer w jego rodzimym środowisku
Używając kombinacji starannie znakowanych próbek i wielowymiarowych eksperymentów NMR, autorzy zbudowali strukturę 3D o wysokiej rozdzielczości dimeru transportera wewnątrz dwuwarstwy lipidowej. Potwierdzili, że białko rzeczywiście tworzy zespół parowy z jamą otwartą na zewnątrz komórki, odpowiednią do wejścia cukru. Gdy zmienili trzy dodatnio naładowane reszty aminokwasowe na interfejsie dimeru, które kontaktują lipidy, struktura parowa stała się krucha: w żelach rozpadała się łatwiej, a jej sygnały NMR ulegały poszerzeniu i zanikowi, co wskazywało na elastyczny, niestabilny kompleks. Symulacje komputerowe potwierdziły ten obraz, pokazując, że bez lipidów interfejsowych dwaj partnerzy oddalają się i tracą wiele stabilizujących kontaktów.
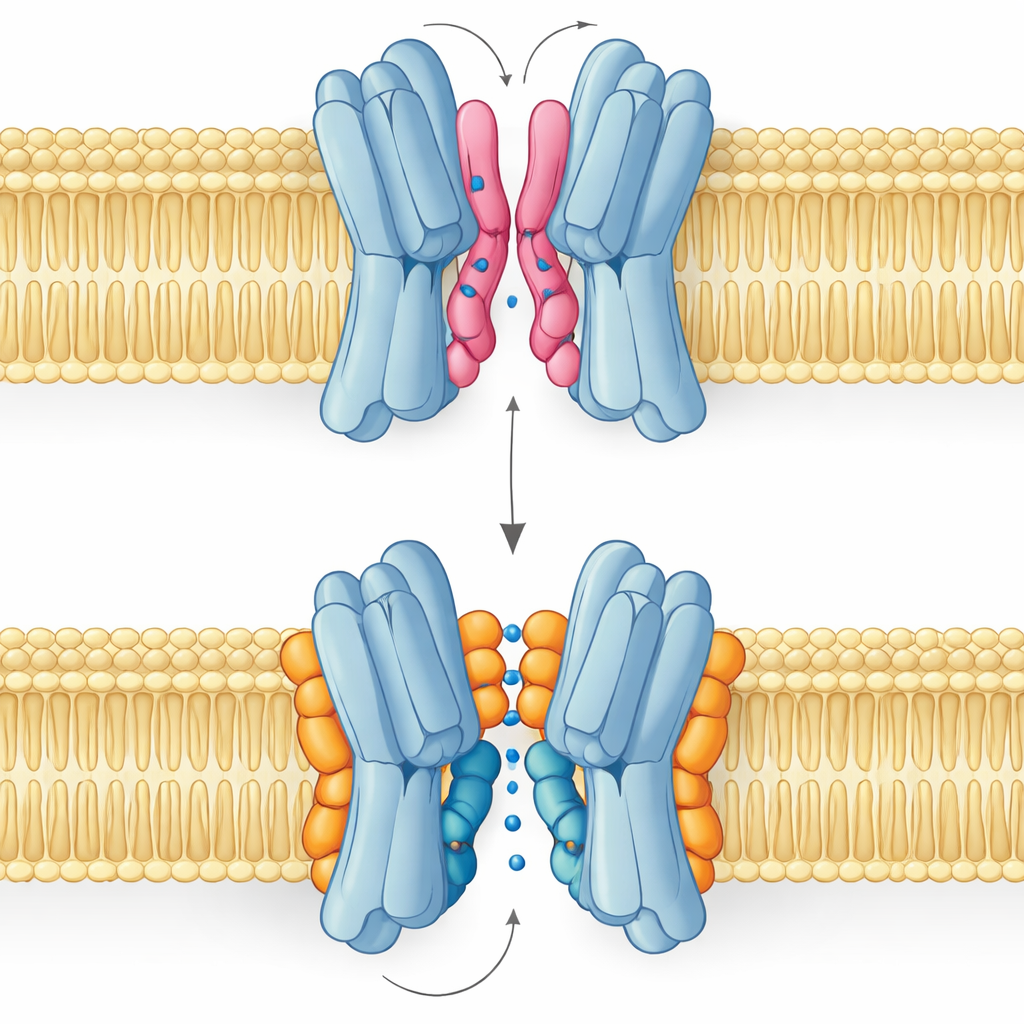
Dobry lipid, zły lipid: opowieść o dwóch trybach wiązania
Najbardziej zaskakujący wynik jest taki, że nie wszystkie lipidy interfejsu pomagają w ten sam sposób. Symulacje i pomiary NMR wykazały, że PG i kardiolipina wiążą się w nakładających się miejscach, ale przyjmują odmienne układy. PG, z dwoma łańcuchami i pojedynczym ładunkiem ujemnym, potrafi wślizgnąć swoje ogony głęboko w wąską szczelinę między partnerami białkowymi, lekko je rozchylając, a nawet zatykać centralne przejście. Kardiolipina natomiast ma masywniejszą główkę niosącą dwa ładunki ujemne i cztery łańcuchy. Jej główka zaciska się na skupisku zasadowych reszt jak zacisk, podczas gdy większe zgrupowanie ogonów pozostaje na zewnętrznej powierzchni, omijając centralną jamę i zamiast tego podpierając jedną z kluczowych helis, która kołysze się podczas cyklu transportowego.
Od wiązania lipidów do transportu cukru
Te różnice strukturalne przekładają się bezpośrednio na funkcję. Widoki energii swobodnej z długich symulacji pokazują, że gdy kardiolipina jest związana, transporter może płynnie odwiedzać konfiguracje otwarte na zewnątrz, zablokowane i otwarte do wnętrza, niezbędne do przeniesienia cukru przez błonę. Najstabilniejsze stany są skierowane na zewnątrz, zgodnie ze strukturą eksperymentalną, ale ścieżka do form skierowanych do wnętrza pozostaje dostępna. Gdy dominuje PG, obraz się zmienia: jej ogony wypełniają centralną jamę, kontakty między podjednostkami ulegają rozluźnieniu, a białko zostaje uwięzione w nieproduktywnej konformacji, przez którą cukier nie może przejść. PE zachowuje się podobnie do PG — sprzyja parowaniu podjednostek, ale czyni je bardziej heterogennymi i mniej stabilnymi niż w stanie związanym z kardiolipiną.
Dlaczego to ma znaczenie poza jednym bakterio-transporterem
Mówiąc prostszym językiem: praca ta pokazuje, że lipidy błonowe nie są tylko biernym smarem, lecz aktywnymi elementami mechanicznymi, które mogą przyczepiać się do białek, podpierać ruchome części albo klinować się w newralgicznych szczelinach. Dla VsSemiSWEET kardiolipina działa jak dopasowana podkładka, która zarówno blokuje dwie połowy razem, jak i ukierunkowuje ruchy potrzebne do transportu cukru, podczas gdy PG ma skłonność do zatykania mechanizmu. Ponieważ wiele białek błonowych ma podobne miejsca interfejsowe i często polega na kardiolipinie lub pokrewnych lipidach, te wyniki stanowią szerszy schemat, jak zmiana składu lipidów może stroić aktywność transporterów, kanałów i pomp w całej biologii.
Cytowanie: Zhang, Y., Zhao, W., Duan, M. et al. Diverse regulation of functional dimerization of a sugar transporter by different interfacial lipids. Nat Commun 17, 3062 (2026). https://doi.org/10.1038/s41467-026-69804-3
Słowa kluczowe: lipidy błonowe, transportery cukrów, kardiolipina, dymeryzacja białek, NMR ciała stałego