Clear Sky Science · sv
Mänsklig FUS är giftig via samspel med RNA-polymeras II i Drosophila
Varför detta är viktigt för hjärnhälsa
ALS (Lou Gehrigs sjukdom) och vissa former av demens förstör nervceller långsamt, men vi förstår fortfarande inte fullt ut varför dessa celler dör. Denna studie använder bananflugor för att undersöka ett protein kallat FUS, vilket är kopplat till vissa ärftliga ALS-fall och även förekommer i klumpar i en demensform som kallas frontotemporal lobär degeneration (FTLD). Genom att fråga var i cellen FUS gör mest skada och vilka partners det interagerar med, avslöjar forskarna en överraskande nukleär väg till toxicitet som kan bidra till att förklara varför neuroner fallerar vid specifika hjärnsjukdomar.
Från flugnerv till mänsklig sjukdom
FUS är ett protein som normalt finns i cellkärnan, där DNA lagras och gener avläses. Hos patienter med FUS-kopplad ALS förflyttar sig muterat FUS ofta ut i omgivande cytoplasma och bildar synliga klumpar. En vanlig idé har varit att dessa cytoplasmatiska depositioner är det som dödar neuroner. I bananflugor räcker det dock i många fall att uttrycka extra normal mänsklig FUS i nervceller för att hämma korrekt utveckling, förkorta livslängd och skada rörelseförmåga. Det antyder att för mycket FUS, även utan mutation, är farligt och ger ett kontrollerat system för att reda ut hur och var faran uppstår.
En oväntad vändning i berättelsen om felplacerade proteiner
Forskarna konstruerade flugor som producerar en version av FUS utan dess inbyggda nukleära "postkod", den så kallade nukleära lokaliseringssekvensen. Utan denna tagg lyckas FUS i hög grad inte komma in i kärnan och ackumuleras istället i cytoplasman. I motsats till förväntningarna var dessa flugor mindre sjuka än flugor som gjorde normal FUS: de överlevde bättre och utvecklades mer normalt, även när proteinmängderna justerades så att de två versionerna fanns i liknande nivåer. Detaljerad avbildning av larv- och vuxenneuroner bekräftade att standard-FUS främst samlades i kärnan, medan den förändrade versionen stannade mestadels utanför. Dessa jämförelser ledde författarna till slutsatsen att FUS i den här modellen skadar neuroner främst genom sin verksamhet i kärnan, inte genom cytoplasmatiska klumpar.
Dolda klumpar i cellens kontrollrum
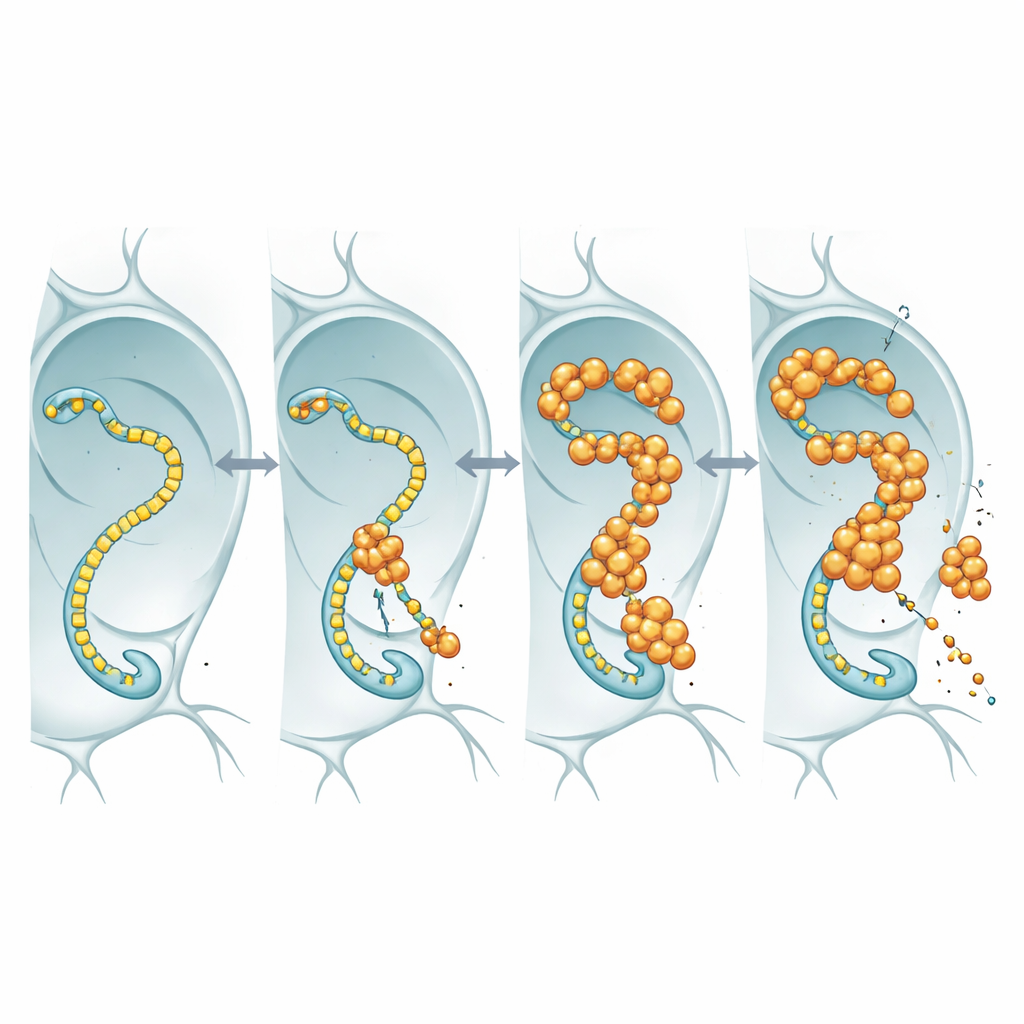
För att se vad nukleärt FUS faktiskt gjorde märkte forskarna FUS med en fluorescerande markör och följde det i de förstorade kärnorna i flugornas spottkörtlar och i neuroner. Istället för att bilda hårda, olösliga aggregat samlades FUS i ljusa, punktformiga granuler som förblev dynamiska: när en fläck blektes med en laser strömmade oblekta molekyler tillbaka inom sekunder. Dessa droppar tenderade att uppträda i regioner som också innehöll höga nivåer av RNA-polymeras II, enzymet som färdas längs DNA för att göra RNA-kopior av gener. Denna samexistens antydde att FUS kan störa den centrala transkriptionsmaskineriet som förser neuronerna med de RNA-meddelanden de behöver.
När genläsarmaskinen blir en medbrottsling
RNA-polymeras II har en flexibel svans bestående av upprepade korta motiv; denna svans fungerar som landningsbana för många reglerande proteiner, inklusive FUS och dess släktingar. Genom att använda fluglinjer där denna svans var konstruerad att innehålla färre eller fler upprepningar undersökte teamet om ändrad svanslängd skulle påverka FUS-toxisk. I flugor som överproducerade FUS gjorde förkortning av svansen att djuren levde längre, även om FUS-RNA och proteinnivåer förblev liknande. Denna längdberoende överensstämmer med tidigare biokemiska studier som visat att längre svansar binder FUS mer entusiastiskt. Den enklaste tolkningen är att FUS utövar sin toxiska effekt genom att engagera denna svans: när svansen är lång binder FUS starkare, rubbar normal transkriptionskontroll och neuroner lider; när svansen förkortas finns det mindre för FUS att gripa tag i och det blir mindre skadligt.
Ledtrådar från mänsklig hjärnvävnad
Författarna vände sig sedan till mänskliga post mortem-prover. I ryggmärgsvävnad från ALS-patienter med FUS-mutationer såg de de förväntade FUS-positiva inklusionerna i neuroner, men den mänskliga versionen av enzymets stora subenhet (POLR2A) förblev i sin vanliga nukleära position. Däremot såg man i frontal cortex från patienter med FTLD kännetecknad av FUS-positiva inklusioner att POLR2A ofta uppträdde i onormala cytoplasmatiska fläckar som ibland överlappade med FUS-ansamlingar. Dessa felplaceringar saknades i kontrollhjärnor. Detta mönster tyder på att i FTLD, men inte nödvändigtvis i FUS-ALS, kan FUS dra med sig RNA-polymeras II ur läge och potentiellt slå ut genuttrycket i drabbade neuroner.
Vad detta betyder för framtida terapier
Tillsammans talar arbetet för att i dessa flugmodeller är den farligaste formen av FUS inte den cytoplasmatiska klumpen utan den nukleära versionen som interagerar med genläsningsmaskineriet. Genom att bilda dynamiska granuler nära RNA-polymeras II och binda dess flexibla svans verkar överskott av FUS störa flödet av genetisk information som behövs för att hålla neuroner vid liv. Observationen att POLR2A också fellokaliseras i mänsklig FTLD-hjärnvävnad stärker argumentet att detta nukleära partnerskap bidrar till sjukdomen. Terapier som begränsar FUS-ansamling i kärnan eller försvagar dess grepp om RNA-polymeras II skulle därför kunna skydda neuroner och erbjuda en ny angreppsvinkel mot ALS-relaterade och FUS-positiva demenser.
Citering: Moens, T.G., Biasetti, L., Scheveneels, W. et al. Human FUS is toxic via association with RNA polymerase II in Drosophila. Cell Death Dis 17, 310 (2026). https://doi.org/10.1038/s41419-026-08539-x
Nyckelord: FUS-protein, RNA-polymeras II, ALS, frontotemporal demens, Drosophila-modell