Clear Sky Science · pt
FUS humano é tóxico por associação com a RNA polimerase II em Drosophila
Por que isso importa para a saúde do cérebro
ELA (doença de Lou Gehrig) e certas formas de demência destroem lentamente neurônios, mas ainda não entendemos completamente por que essas células morrem. Este estudo usa moscas-das-frutas para investigar uma proteína chamada FUS, que está associada a alguns casos hereditários de ELA e também aparece em agregados numa forma de demência chamada degeneração lobar frontotemporal (DLFT). Ao perguntar onde, dentro da célula, o FUS causa mais dano e com quais parceiros interage, os pesquisadores revelam uma via nuclear surpreendente para a toxicidade que pode ajudar a explicar por que neurônios falham em doenças cerebrais específicas.
Das fibras nervosas da mosca para a doença humana
O FUS é uma proteína que normalmente reside no núcleo celular, onde o DNA é armazenado e os genes são lidos. Em pacientes com ELA associada a FUS, o FUS mutante frequentemente se desloca para o citoplasma circundante, formando agregados visíveis. Uma ideia popular tem sido que esses depósitos citoplasmáticos são os responsáveis pela morte neuronal. Em moscas-das-frutas, entretanto, simplesmente produzir excesso de FUS humano normal nas células nervosas já é suficiente para bloquear o desenvolvimento adequado, encurtar a vida e prejudicar o movimento. Isso sugere que ter FUS em excesso, mesmo sem mutação, é perigoso e fornece um sistema controlado para separar como e onde esse perigo surge.
Uma reviravolta na história do deslocamento proteico
A equipe engenheirou moscas para produzir uma versão do FUS que carece de seu “código postal” nuclear embutido, conhecido como sequência de localização nuclear. Sem essa etiqueta, o FUS em grande parte deixa de entrar no núcleo e, em vez disso, acumula-se no citoplasma. Contrariando as expectativas, essas moscas eram menos doentes do que moscas produzindo FUS normal: sobreviviam melhor e se desenvolviam mais normalmente, mesmo quando os níveis de proteína foram ajustados para que as duas versões estivessem presentes em quantidades semelhantes. Imagens detalhadas de neurônios larvais e adultos confirmaram que o FUS padrão se acumulava principalmente no núcleo, enquanto a versão alterada permanecia majoritariamente fora. Essas comparações levaram os autores a concluir que, nesse modelo, o FUS prejudica os neurônios principalmente pelo que faz no núcleo, e não por agregados citoplasmáticos.
Agressados ocultos na sala de controle da célula
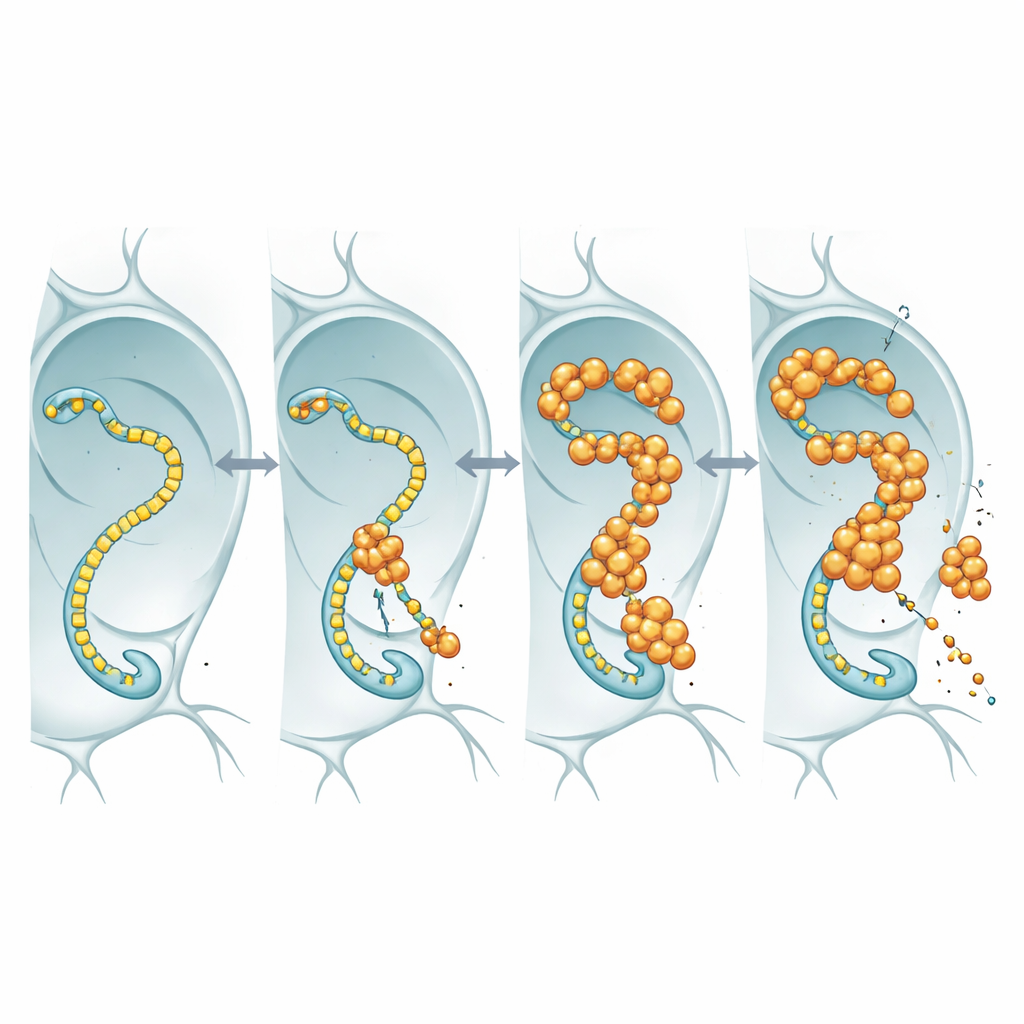
Para ver o que o FUS nuclear está realmente fazendo, os pesquisadores marcaram o FUS com um marcador fluorescente e o observaram nos núcleos sobredimensionados das glândulas salivares da mosca e em neurônios. Em vez de formar agregados duros e insolúveis, o FUS reunia-se em grânulos pontuais brilhantes que permaneciam dinâmicos: quando uma região era branqueada com um laser, moléculas não branqueadas fluíam de volta em questão de segundos. Essas gotículas tendiam a aparecer em regiões que também continham altos níveis de RNA polimerase II, a enzima que percorre o DNA para fabricar cópias de RNA dos genes. Essa colocalização sugeriu que o FUS poderia estar interferindo na maquinaria central de transcrição que mantém os neurônios supridos com as mensagens em RNA de que precisam.
Quando a máquina de ler genes vira cúmplice
A RNA polimerase II possui uma cauda flexível composta por motivos curtos repetidos; essa cauda atua como uma pista de pouso para muitas proteínas regulatórias, incluindo o FUS e seus parentes. Usando linhagens de mosca nas quais essa cauda foi projetada para conter menos ou mais repetições, a equipe investigou se alterar o comprimento da cauda mudaria a toxicidade do FUS. Em moscas que superproduziam FUS, encurtar a cauda fez os animais viverem mais, mesmo que os níveis de RNA e proteína de FUS permanecessem similares. Essa dependência do comprimento corresponde a trabalhos bioquímicos anteriores que mostram que caudas mais longas ligam-se ao FUS com maior afinidade. A interpretação mais simples é que o FUS exerce seu efeito tóxico ao engajar essa cauda: quando a cauda é longa, o FUS se liga mais fortemente, perturba o controle transcricional normal e os neurônios sofrem; quando a cauda é encurtada, o FUS tem menos onde se agarrar e torna-se menos nocivo.
Pistas de tecido cerebral humano
Os autores então recorreram a amostras humanas pós-morte. Em tecido de medula espinhal de pacientes com ELA portadores de mutações em FUS, eles observaram as esperadas inclusões positivas para FUS em neurônios, mas a versão humana da subunidade grande da enzima (POLR2A) permaneceu em sua localização nuclear usual. Em contraste, no córtex frontal de pacientes com DLFT marcada por inclusões positivas para FUS, a POLR2A frequentemente aparecia em pontos citoplasmáticos anormais que às vezes se sobrepunham aos depósitos de FUS. Esses deslocamentos estavam ausentes em cérebros-controle. Esse padrão sugere que, na DLFT, mas não necessariamente na ELA associada a FUS, o FUS pode arrastar a RNA polimerase II para fora de posição, potencialmente comprometendo a expressão gênica nos neurônios afetados.
O que isso significa para terapias futuras
Em conjunto, o trabalho indica que, nesses modelos em mosca, a forma mais perigosa de FUS não é o agregado citoplasmático, mas a versão nuclear que interage com a maquinaria de leitura dos genes. Ao formar grânulos dinâmicos próximos à RNA polimerase II e ligar-se à sua cauda flexível, o excesso de FUS parece perturbar o fluxo de informação genética necessário para manter os neurônios vivos. A observação de que a POLR2A também se deslocaliza em tecido cerebral humano de DLFT fortalece a hipótese de que essa parceria nuclear contribui para a doença. Terapias que limitem o acúmulo de FUS no núcleo ou enfraqueçam sua ligação à RNA polimerase II podem, portanto, proteger neurônios, oferecendo um novo ângulo de ataque contra ELA relacionada e demências positivas para FUS.
Citação: Moens, T.G., Biasetti, L., Scheveneels, W. et al. Human FUS is toxic via association with RNA polymerase II in Drosophila. Cell Death Dis 17, 310 (2026). https://doi.org/10.1038/s41419-026-08539-x
Palavras-chave: Proteína FUS, RNA polimerase II, ELA, demência frontotemporal, modelo em Drosophila