Clear Sky Science · ru
Молекулярная основа селективности силы у PIEZO2
Как наши нервы отличают укол от растяжения
Каждый раз, когда вы проводите кончиками пальцев по странице или чувствуете, как при вдохе расширяются лёгкие, крошечные молекулярные машины в клетках преобразуют механическую силу в электрические сигналы. Два близкородственных канала, называемые PIEZO1 и PIEZO2, лежат в основе этого процесса. И всё же они реагируют на очень разные виды сил: один предпочитает широкое растяжение, другой «заточён» под лёгкие уколы и надавливания, которые мы воспринимаем как прикосновение. Эта работа раскрывает, как PIEZO2 достигает такой специализации, показывая физический «тросик», который помогает нашей нервной системе различать виды механических воздействий.
Два «сиблинг»-сенсора с разными задачами
PIEZO1 и PIEZO2 — крупные белки в форме трипода, расположенные в клеточной мембране, которые открываются при деформации мембраны. PIEZO1 в основном встречается в непроводящих нерв клетках, таких как эритроциты и хрящ, где он хорошо реагирует на общее растяжение или поток жидкости. PIEZO2, напротив, сконцентрирован в нейронах, отвечающих за осязание и положение тела, и необходим для лёгкого прикосновения, проприоцепции и дыхания. При взгляде под микроскопом их структуры выглядят поразительно похоже, что породило вопрос: почему PIEZO1 сильно реагирует на растяжение мембраны, тогда как PIEZO2 гораздо чувствительнее к локальной индукции, как маленький укол по поверхности клетки.
Наблюдая за движением одиночных молекул под действием силы
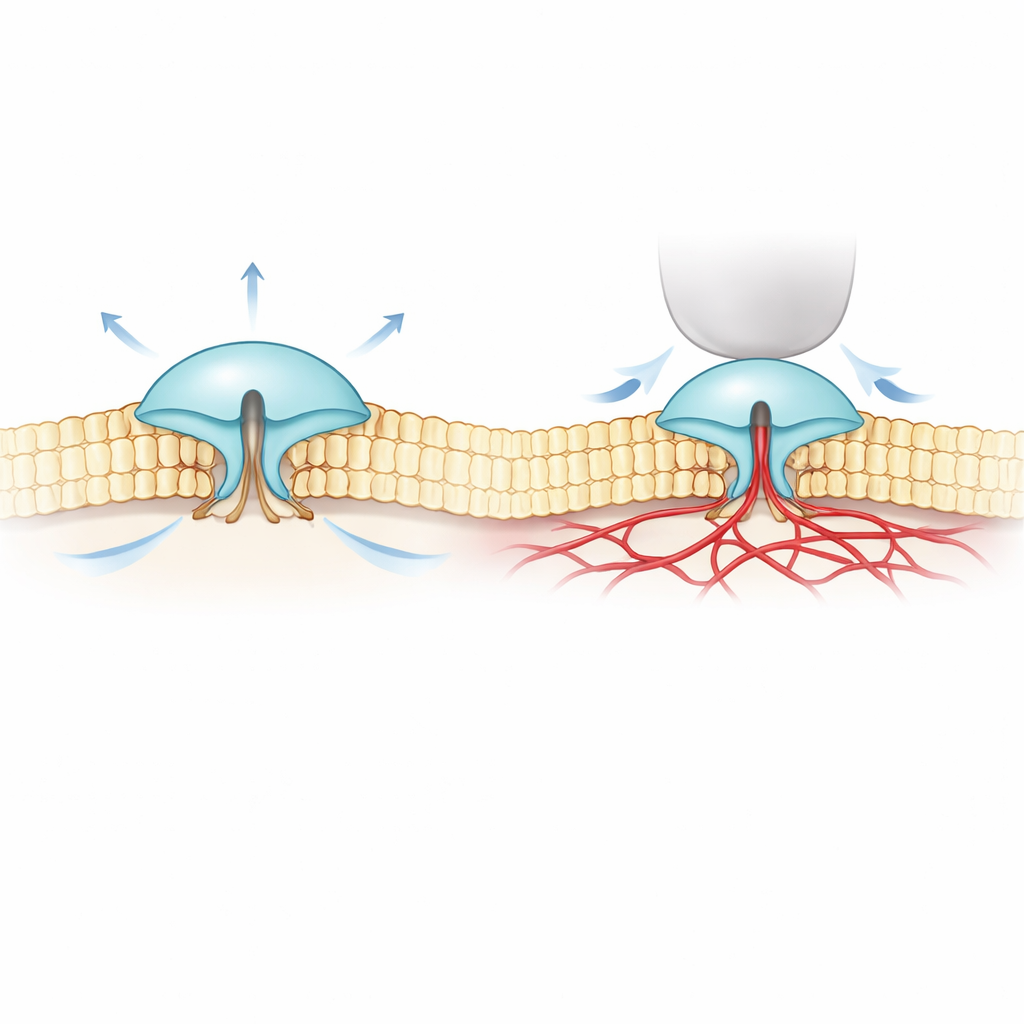
Чтобы разобраться в этом, авторы объединили сверхточную флуоресцентную микроскопию (MINFLUX) с электрическими записями. Они создали версии PIEZO1 и PIEZO2 с маленькими светочувствительными метками на внешних частях их «лопастей» и следили за изменением расстояния между этими лопастями в настоящих клеточных мембранах. Они обнаружили, что PIEZO2 по своей сути более компактный и жёсткий, чем PIEZO1: его лопасти меньше флуктуируют и находятся ближе друг к другу в состоянии покоя. Когда клетки раздували, чтобы увеличить натяжение мембраны — аналогично надуванию шарика — лопасти PIEZO1 расходились, и канал открывался, что соответствует простой модели «открывается под тягой мембраны». PIEZO2 вёл себя противоположно: его лопасти слегка сжимались, а канал едва открывался, несмотря на общее натяжение мембраны.
Скрытая связь с внутренним каркасом клетки
Эти неожиданные результаты предположили, что PIEZO2 управляется не столько окружающим жировым слоем мембраны, сколько чем‑то внутри клетки. Команда обратила внимание на актиновый цитоскелет — сеть белковых филаментов прямо под мембраной, придающую клетке форму. В клеточном типе с высоким содержанием актина уменьшение объёма клетки (что ослабляет мембрану и складирует её в актиновой сети) фактически вызывало расширение лопастей PIEZO2, снова в противоположность PIEZO1. Когда исследователи химически разрушали актин или удаляли гибкий внутренний сегмент PIEZO2, ранее связанный с осязанием, поведение PIEZO2 менялось: он начинал реагировать на растяжение гораздо как PIEZO1. Треккинг одиночных молекул также показал, что нормальный PIEZO2 почти не перемещается в мембране — что соответствует его закреплению — тогда как после удаления этого внутреннего сегмента он свободно диффундирует.
Filamin‑B: тросик, настраивающий осязание
Чтобы найти недостающую связь между PIEZO2 и актином, авторы перекрестно сшили близлежащие белки с PIEZO2 и идентифицировали их методом масс‑спектрометрии. Среди нескольких кандидатов выделился filamin‑B (FLNB) — крупный каркасный белок, известный связью с актином и множеством мембранных белков. Подавление FLNB в инженерных клетках превратило PIEZO2 в надёжный сенсор растяжения и увеличило его мобильность в мембране, имитируя эффект удаления внутреннего сегмента. Важно, что удаление FLNB снизило чувствительность PIEZO2 к надавливанию тупым зондом и увеличило глубину вдавления, необходимую для его открытия. В сенсорных нейронах мышей частичная потеря FLNB заставляла родные каналы PIEZO2 реагировать на растяжение мембраны, на которое они обычно не реагируют, подтверждая, что FLNB необходим для их обычного предпочтения индукции.
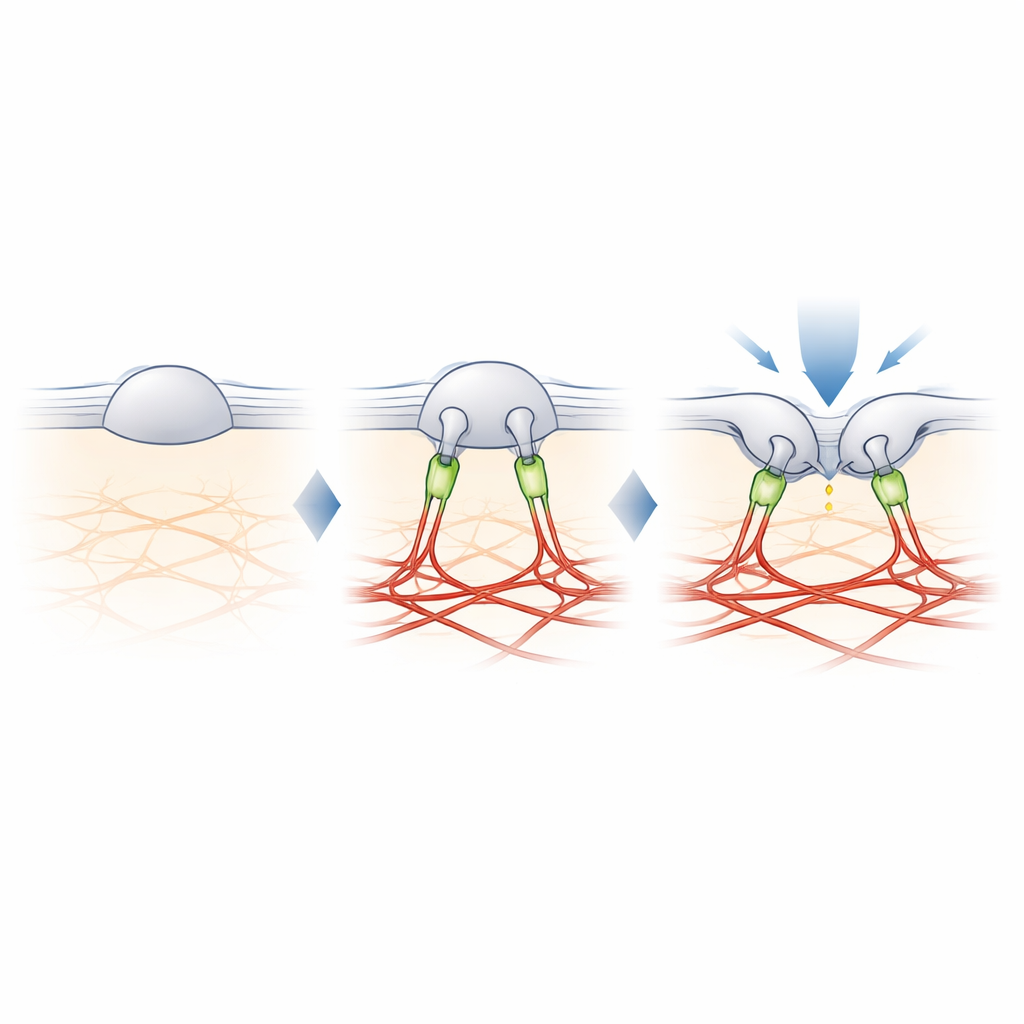
От молекулярного тросика к чувству осязания
Исследование делает вывод, что особая роль PIEZO2 в осязании обусловлена физической привязью к актиновому каркасу, в которой участвует filamin‑B. Вместо того чтобы просто ощущать степень растяжения мембраны, PIEZO2 фиксирует, как мембрана изгибается и смещается относительно этого внутреннего якоря, что поддерживает канал в слегка приоткрытом состоянии и делает его легче возбудимым при локальном уколе. PIEZO1, напротив, остаётся в основном незафиксированным и реагирует главным образом на общее натяжение. Это молекулярное распределение функций помогает объяснить, как наши тела используют близкородственные каналы для различения лёгкого прикосновения и глобальных сил, таких как кровоток или давление. Это также даёт схему того, как клетки в разных тканях могут тонко настраивать свою механическую чувствительность, добавляя или убирая подобные «тросики».
Цитирование: Mulhall, E.M., Yarishkin, O., Hill, R.Z. et al. The molecular basis of force selectivity by PIEZO2. Nature 653, 297–305 (2026). https://doi.org/10.1038/s41586-026-10182-7
Ключевые слова: механосенсация, PIEZO2, механорецепторы прикосновения, цитоскелет, ионные каналы