Clear Sky Science · it
La base molecolare della selettività di forza di PIEZO2
Come i nostri nervi distinguono una pressione da un allungamento
Ogni volta che sfiori le pagine con la punta delle dita o senti i polmoni espandersi mentre respiri, minuscole macchine molecolari nelle cellule convertono la forza meccanica in segnali elettrici. Due canali strettamente correlati, chiamati PIEZO1 e PIEZO2, sono al centro di questo processo. Tuttavia rispondono a tipi di forza molto diversi: uno predilige l’allungamento globale, l’altro è tarato per le carezze e le pressioni leggere che percepiamo come tatto. Questo articolo svela come PIEZO2 ottenga questa specializzazione, rivelando un legame fisico che aiuta il nostro sistema nervoso a distinguere un tipo di forza dall’altro.
Due sensori “fratelli” con compiti diversi
PIEZO1 e PIEZO2 sono grandi proteine a forma di treppiede che si trovano nella membrana cellulare e si aprono quando la membrana viene deformata. PIEZO1 si trova principalmente in cellule non nervose, come i globuli rossi e la cartilagine, dove risponde bene all’allungamento generale o al flusso di liquidi. PIEZO2, invece, è concentrato nei neuroni sensoriali del tatto e della posizione ed è cruciale per il tatto leggero, la propriocezione e la respirazione. Al microscopio le loro strutture appaiono sorprendentemente simili, il che rendeva difficile capire perché PIEZO1 reagisca fortemente allo stiramento della membrana, mentre PIEZO2 risponda molto meglio all’indentazione locale, come un piccolo pizzico sulla superficie cellulare.
Osservare singole molecole muoversi sotto forza
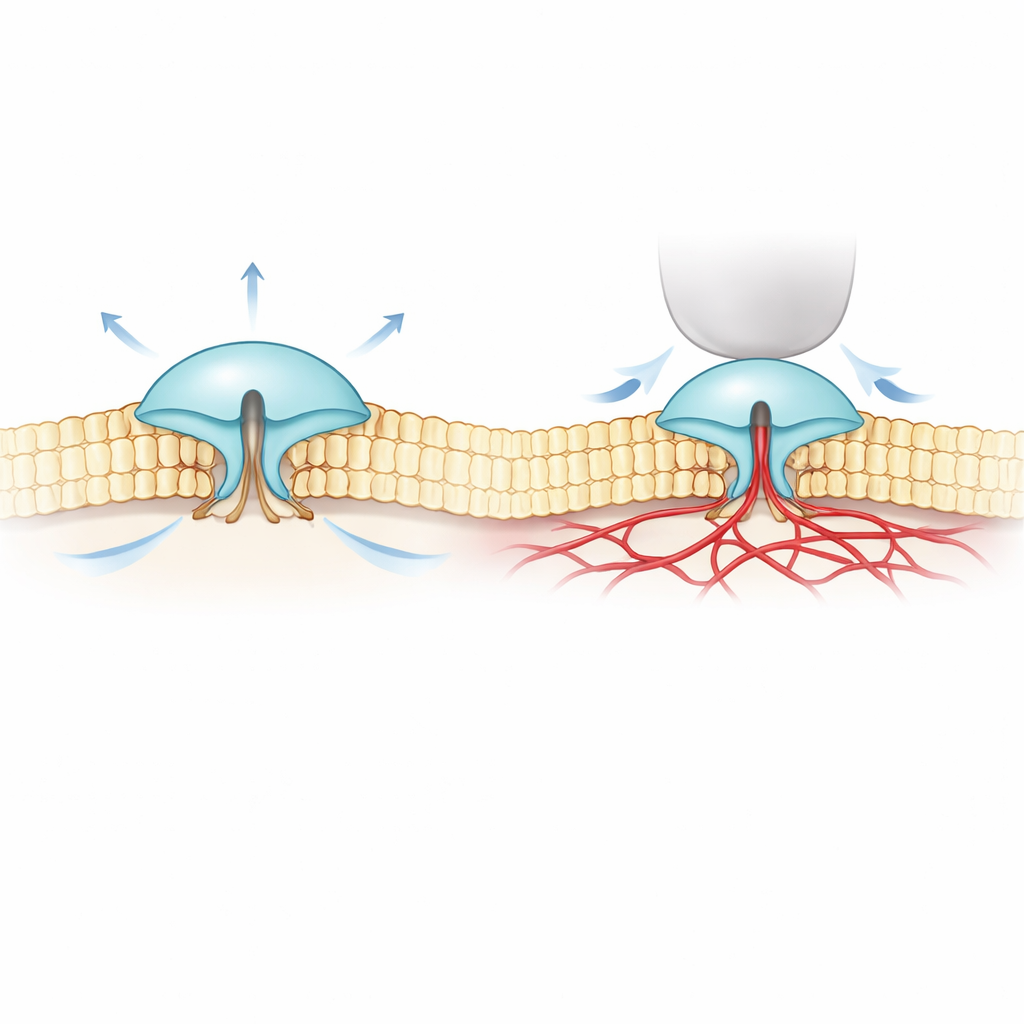
Per affrontare questo mistero, gli autori hanno combinato l’imaging fluorescente ultra-preciso (chiamato MINFLUX) con registrazioni elettriche. Hanno costruito versioni di PIEZO1 e PIEZO2 che portano minuscoli marcatori sensibili alla luce sulle parti esterne delle loro “lame” e hanno osservato come cambiava la distanza tra queste lame nelle membrane cellulari reali. Hanno scoperto che PIEZO2 è intrinsecamente più compatto e rigido rispetto a PIEZO1: le sue lame fluttuano meno e a riposo stanno più vicine. Quando le cellule venivano gonfiate per aumentare la tensione della membrana — simile a infiammare un palloncino — le lame di PIEZO1 si allargavano e il canale si apriva, coerente con un semplice modello di “apertura tirata dalla membrana”. PIEZO2 si comportava in modo opposto: le sue lame si stringevano leggermente e il canale si apriva pochissimo, nonostante l’intera membrana fosse sotto tensione.
Un collegamento nascosto con l’impalcatura interna della cellula
Questi risultati sorprendenti suggerivano che PIEZO2 non sia guidato principalmente dallo strato lipidico circostante della membrana, ma da qualcos’altro all’interno della cellula. Il gruppo si è rivolto al citoscheletro di actina, una rete di filamenti proteici appena sotto la membrana che aiuta a dare forma alle cellule. In un tipo cellulare ricco di actina, ridurre il volume cellulare (che allenta la membrana e la ripiega nella rete di actina) faceva espandere le lame di PIEZO2, di nuovo in modo opposto rispetto a PIEZO1. Quando i ricercatori hanno disturbato chimicamente l’actina, o hanno cancellato un segmento interno flessibile di PIEZO2 precedentemente collegato al tatto, il comportamento di PIEZO2 è cambiato: ha cominciato a rispondere allo stiramento molto più come PIEZO1. Il tracciamento a singola molecola ha inoltre mostrato che il PIEZO2 normale si muove pochissimo nella membrana — coerente con essere ancorato — mentre dopo l’interruzione di quel segmento interno diffonde liberamente.
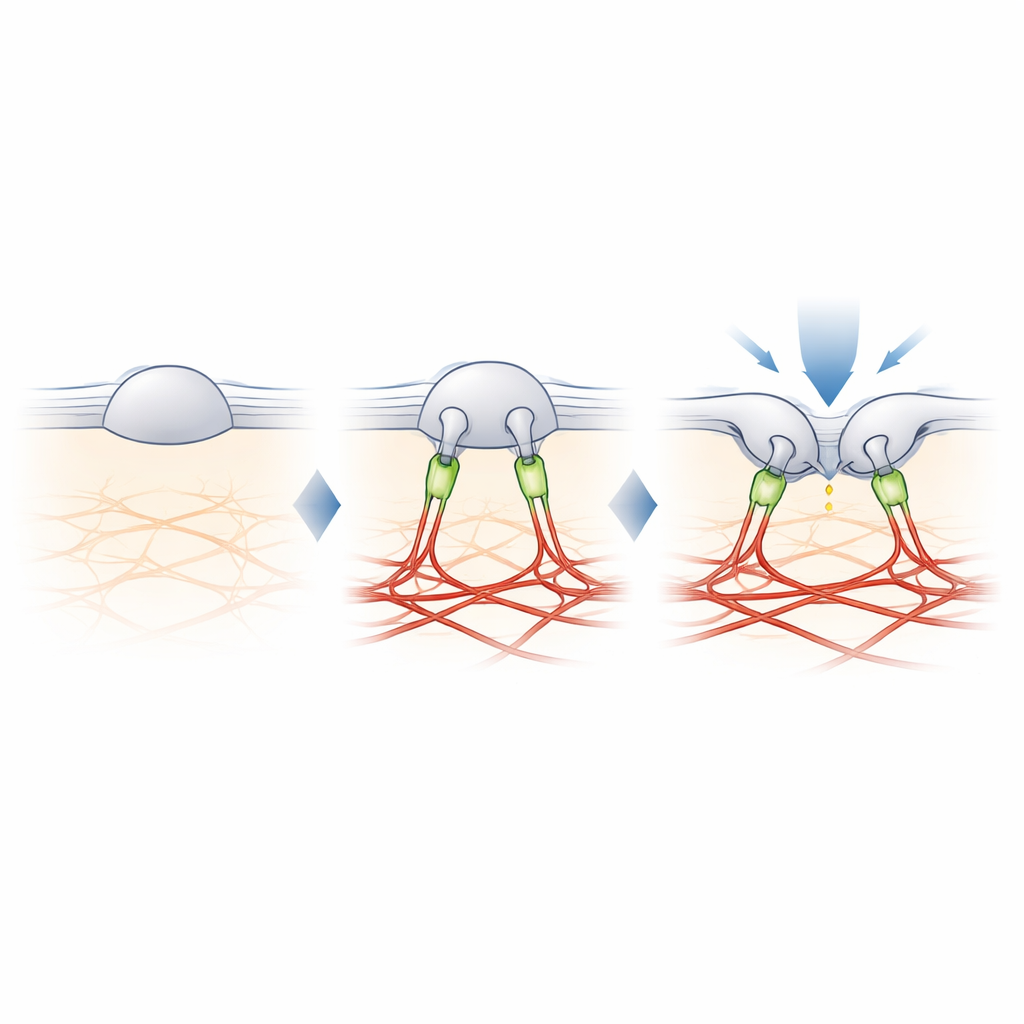
Filamin-B: il legame che modula il tatto
Per trovare il collegamento mancante tra PIEZO2 e l’actina, gli autori hanno cross-legato le proteine vicine a PIEZO2 e le hanno identificate tramite spettrometria di massa. Tra diversi candidati, uno è emerso chiaramente: filamin-B (FLNB), una grande proteina di impalcatura nota per legare l’actina e numerose proteine di membrana. Ridurre l’espressione di FLNB in cellule ingegnerizzate trasformava PIEZO2 in un solido sensore di stiramento e aumentava la sua mobilità nella membrana, imitando gli effetti del taglio del suo segmento interno. Crucialmente, la rimozione di FLNB riduceva la sensibilità di PIEZO2 ai pizzichi con una sonda ottusa e aumentava la profondità di indentazione necessaria per aprirlo. Nei neuroni sensoriali di topo, la perdita parziale di FLNB rendeva i canali PIEZO2 nativi sensibili allo stiramento della membrana che normalmente ignorano, confermando che FLNB è essenziale per la loro abituale preferenza per l’indentazione.
Dal legame molecolare al senso del tatto
Lo studio conclude che il ruolo speciale di PIEZO2 nel tatto deriva da un legame fisico con l’impalcatura di actina, in parte mediato da filamin-B. Piuttosto che misurare semplicemente quanto la membrana è stirata, PIEZO2 percepisce come la membrana si piega e si sposta rispetto a questo ancoraggio interno, che mantiene il canale leggermente pre-aperto e più facile da attivare con un pizzico locale. PIEZO1, al contrario, rimane per lo più non ancorato e risponde principalmente alla tensione globale. Questa divisione molecolare del lavoro aiuta a spiegare come i nostri corpi usino canali strettamente correlati per distinguere il tatto leggero da forze globali come il flusso sanguigno o la pressione. Offre inoltre una guida su come le cellule in molti tessuti possano modulare la loro sensibilità meccanica aggiungendo o rimuovendo tali ancoraggi.
Citazione: Mulhall, E.M., Yarishkin, O., Hill, R.Z. et al. The molecular basis of force selectivity by PIEZO2. Nature 653, 297–305 (2026). https://doi.org/10.1038/s41586-026-10182-7
Parole chiave: meccanosensazione, PIEZO2, neuroni del tatto, cito Scheletro, canali ionici