Clear Sky Science · es
La base molecular de la selectividad de fuerzas por PIEZO2
Cómo nuestros nervios distinguen un pellizco de un estiramiento
Cada vez que deslizas las yemas de los dedos sobre una página o sientes cómo se expanden tus pulmones al respirar, diminutas máquinas moleculares en tus células convierten la fuerza mecánica en señales eléctricas. Dos canales estrechamente relacionados, llamados PIEZO1 y PIEZO2, son centrales en este proceso. Sin embargo, responden a tipos de fuerza muy distintos: uno prefiere el estiramiento amplio, el otro está afinado para toques y presiones suaves que percibimos como tacto. Este artículo revela cómo PIEZO2 logra esa especialización, mostrando un anclaje físico que ayuda a nuestro sistema nervioso a distinguir un tipo de fuerza de otro.
Dos sensores hermanos con trabajos diferentes
PIEZO1 y PIEZO2 son proteínas grandes con forma de trípode que se sitúan en la membrana celular y se abren cuando la membrana se deforma. PIEZO1 se encuentra sobre todo en células no neuronales, como los glóbulos rojos y el cartílago, donde responde bien al estiramiento general o al flujo de fluidos. PIEZO2, en contraste, está concentrado en neuronas sensoriales del tacto y de la posición y es crucial para el tacto suave, la propiocepción y la respiración. Al observarlas al microscopio, sus estructuras parecen notablemente semejantes, lo que ha hecho enigmático por qué PIEZO1 reacciona con fuerza al estiramiento de la membrana, mientras que PIEZO2 responde mucho mejor a la indentación local, como un pequeño dedo que pulsa la superficie celular.
Observando moléculas individuales moverse bajo fuerza
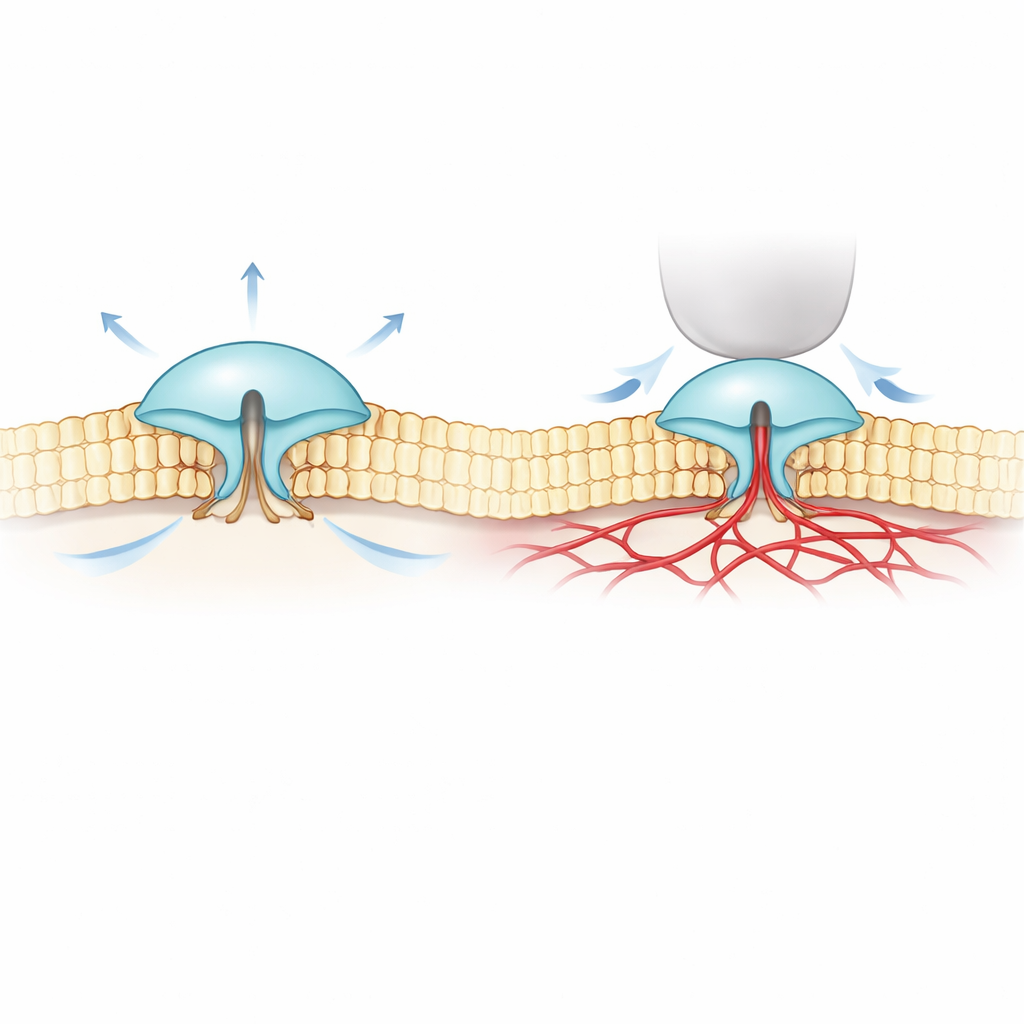
Para abordar este misterio, los autores combinaron imagen fluorescente ultrafina (llamada MINFLUX) con registros eléctricos. Ingeniaron versiones de PIEZO1 y PIEZO2 que llevan pequeñas etiquetas luminiscentes en las partes externas de sus “palas” y observaron cómo cambiaba la separación de esas palas en membranas celulares reales. Encontraron que PIEZO2 es intrínsecamente más compacto y rígido que PIEZO1: sus palas fluctúan menos y están más juntas en reposo. Cuando las células se hincharon para aumentar la tensión de la membrana —similar a inflar un globo— las palas de PIEZO1 se separaron y el canal se abrió, acorde con una imagen simple de “tirado por la membrana”. PIEZO2 se comportó de forma opuesta. Sus palas se apretaron ligeramente y el canal apenas se abrió, pese a que toda la membrana estaba bajo tensión.
Una conexión oculta con el andamiaje interno de la célula
Estos resultados sorprendentes sugirieron que PIEZO2 no está impulsado principalmente por la capa lipídica circundante de la membrana, sino por algo dentro de la célula. El equipo dirigió su atención al citoesqueleto de actina, una malla de filamentos proteicos justo bajo la membrana que ayuda a dar forma a las células. En un tipo celular rico en actina, reducir el volumen celular (lo que afloja la membrana y la pliega sobre la malla de actina) en realidad provocó que las palas de PIEZO2 se expandieran, de nuevo en sentido contrario a PIEZO1. Cuando los investigadores desgarraron químicamente la actina, o eliminaron un segmento interno flexible de PIEZO2 previamente vinculado al tacto, el comportamiento de PIEZO2 se invirtió: comenzó a responder al estiramiento más parecido a PIEZO1. El seguimiento de moléculas individuales mostró además que PIEZO2 normal se mueve muy poco en la membrana —coherente con estar anclado— mientras que tras eliminar ese segmento interno difunde libremente.
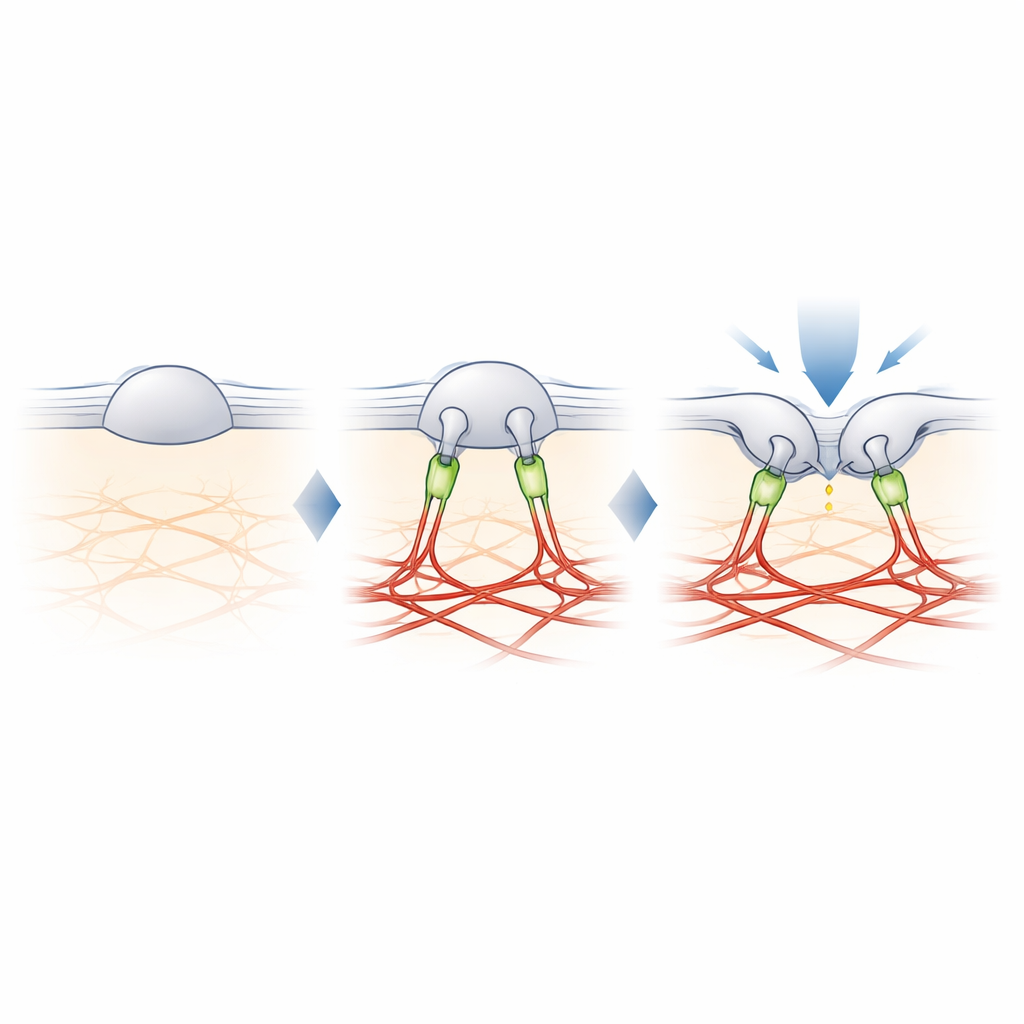
Filamina-B: el cable que afina el tacto
Para encontrar el eslabón perdido entre PIEZO2 y la actina, los autores entrecruzaron proteínas cercanas a PIEZO2 y las identificaron mediante espectrometría de masas. Entre varios candidatos destacó uno: filamina-B (FLNB), una gran proteína andamiaje conocida por unirse a la actina y a muchas proteínas de membrana. Reducir FLNB en células modificadas convirtió a PIEZO2 en un sensor de estiramiento robusto y aumentó su movilidad en la membrana, imitando los efectos de eliminar su segmento interno. De forma crucial, eliminar FLNB redujo la sensibilidad de PIEZO2 a ser pinchado con una sonda roma y elevó la indentación necesaria para abrirlo. En neuronas sensoriales de ratón, la pérdida parcial de FLNB hizo que los canales PIEZO2 nativos respondieran al estiramiento de membrana que normalmente ignoran, confirmando que FLNB es esencial para su preferencia habitual por la indentación.
Del anclaje molecular al sentido del tacto
El estudio concluye que el papel especial de PIEZO2 en el tacto proviene de un anclaje físico al andamiaje de actina, construido en parte por la filamina-B. En lugar de sentir únicamente cuánto se estira la membrana, PIEZO2 percibe cómo la membrana se dobla y se desplaza respecto a este anclaje interno, lo que mantiene el canal ligeramente predispuesto a abrirse y más fácil de activar por una presión local. PIEZO1, en cambio, permanece mayormente sin anclar y responde principalmente a la tensión global. Esta división molecular del trabajo ayuda a explicar cómo nuestros cuerpos usan canales estrechamente relacionados para distinguir el tacto suave de fuerzas globales como el flujo sanguíneo o la presión. También ofrece un modelo de cómo las células en muchos tejidos podrían ajustar su sensibilidad mecánica añadiendo o quitando tales anclajes.
Cita: Mulhall, E.M., Yarishkin, O., Hill, R.Z. et al. The molecular basis of force selectivity by PIEZO2. Nature 653, 297–305 (2026). https://doi.org/10.1038/s41586-026-10182-7
Palabras clave: mecanosensación, PIEZO2, neuronas del tacto, citoesqueleto, canales iónicos