Clear Sky Science · en
The molecular basis of force selectivity by PIEZO2
How Our Nerves Tell a Poke from a Stretch
Every time you brush your fingertips over a page or feel your lungs expand as you breathe, tiny molecular machines in your cells convert mechanical force into electrical signals. Two closely related channels, called PIEZO1 and PIEZO2, are central to this process. Yet they respond to very different kinds of forces: one prefers broad stretching, the other is tuned to gentle pokes and presses that we perceive as touch. This paper uncovers how PIEZO2 pulls off that specialization, revealing a physical tether that helps our nervous system tell one kind of force from another.
Two Sibling Sensors with Different Jobs
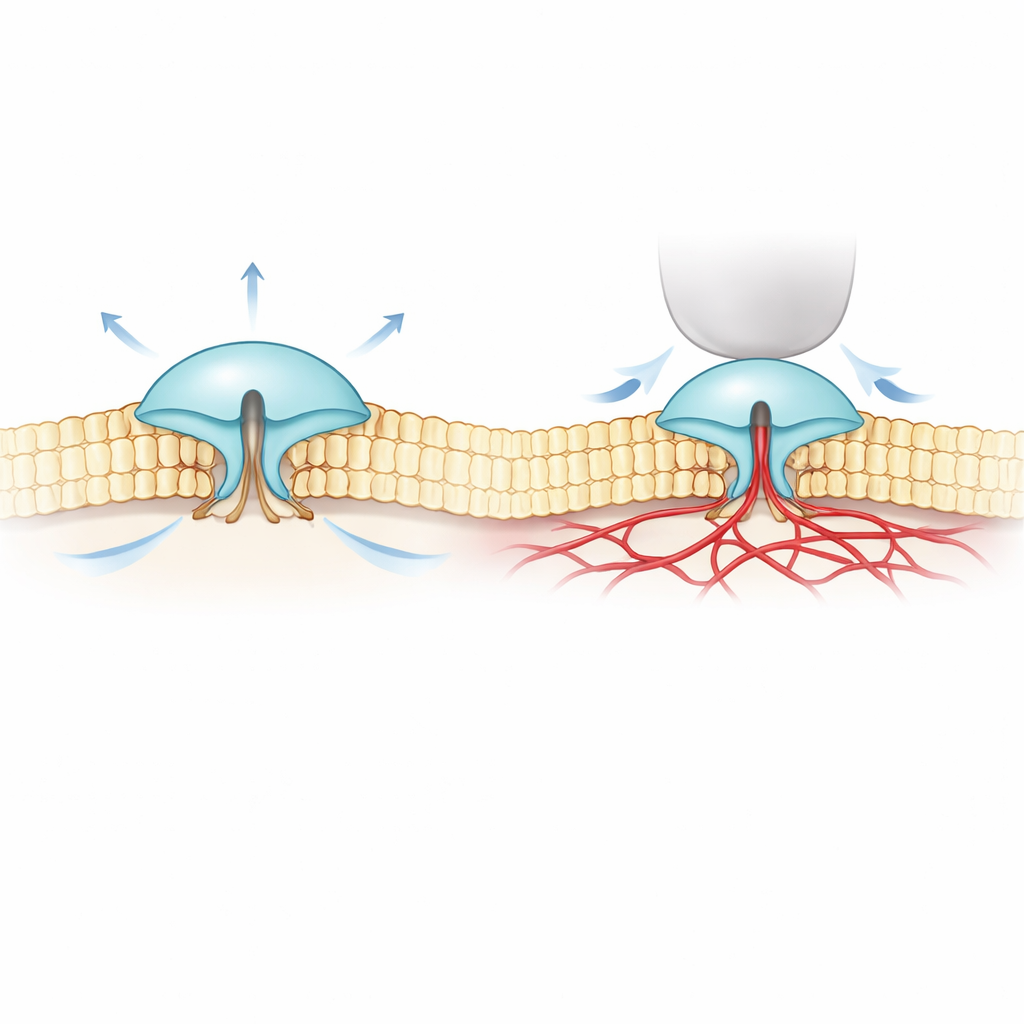
PIEZO1 and PIEZO2 are large, tripod-shaped proteins that sit in the cell membrane and open when the membrane is deformed. PIEZO1 is found mainly in non-nerve cells such as red blood cells and cartilage, where it responds well to overall stretching or fluid flow. PIEZO2, in contrast, is concentrated in touch and position-sensing neurons and is crucial for gentle touch, body awareness, and breathing. Under the microscope their structures look strikingly similar, which has made it puzzling why PIEZO1 reacts strongly to membrane stretch, while PIEZO2 responds far better to local indentation, like a tiny finger poke on the cell surface.
Watching Single Molecules Move Under Force
To tackle this mystery, the authors combined ultra-precise fluorescence imaging (called MINFLUX) with electrical recordings. They engineered versions of PIEZO1 and PIEZO2 that carry tiny light-responsive tags on the outer parts of their “blades” and watched how the spacing of these blades changed in real cell membranes. They found that PIEZO2 is intrinsically more compact and rigid than PIEZO1: its blades fluctuate less and sit closer together at rest. When cells were swollen to increase membrane tension—similar to inflating a balloon—PIEZO1’s blades spread apart and the channel opened, matching a simple “pulled open by the membrane” picture. PIEZO2 behaved in the opposite way. Its blades tightened slightly and the channel barely opened, even though the entire membrane was under tension.
A Hidden Connection to the Cell’s Inner Scaffold
These surprising results suggested that PIEZO2 is not driven mainly by the surrounding fat layer of the membrane but by something else inside the cell. The team looked to the actin cytoskeleton, a mesh of protein filaments just under the membrane that helps give cells their shape. In a concentrated actin-rich cell type, reducing cell volume (which slackens the membrane and folds it into the actin mesh) actually caused PIEZO2’s blades to expand, again opposite to PIEZO1. When the researchers chemically disrupted actin, or deleted a floppy internal segment of PIEZO2 previously linked to touch, PIEZO2’s behavior flipped: now it started to respond to stretch much more like PIEZO1. Single-molecule tracking further showed that normal PIEZO2 hardly moves in the membrane—consistent with being anchored—whereas after disrupting this internal segment it diffuses freely.
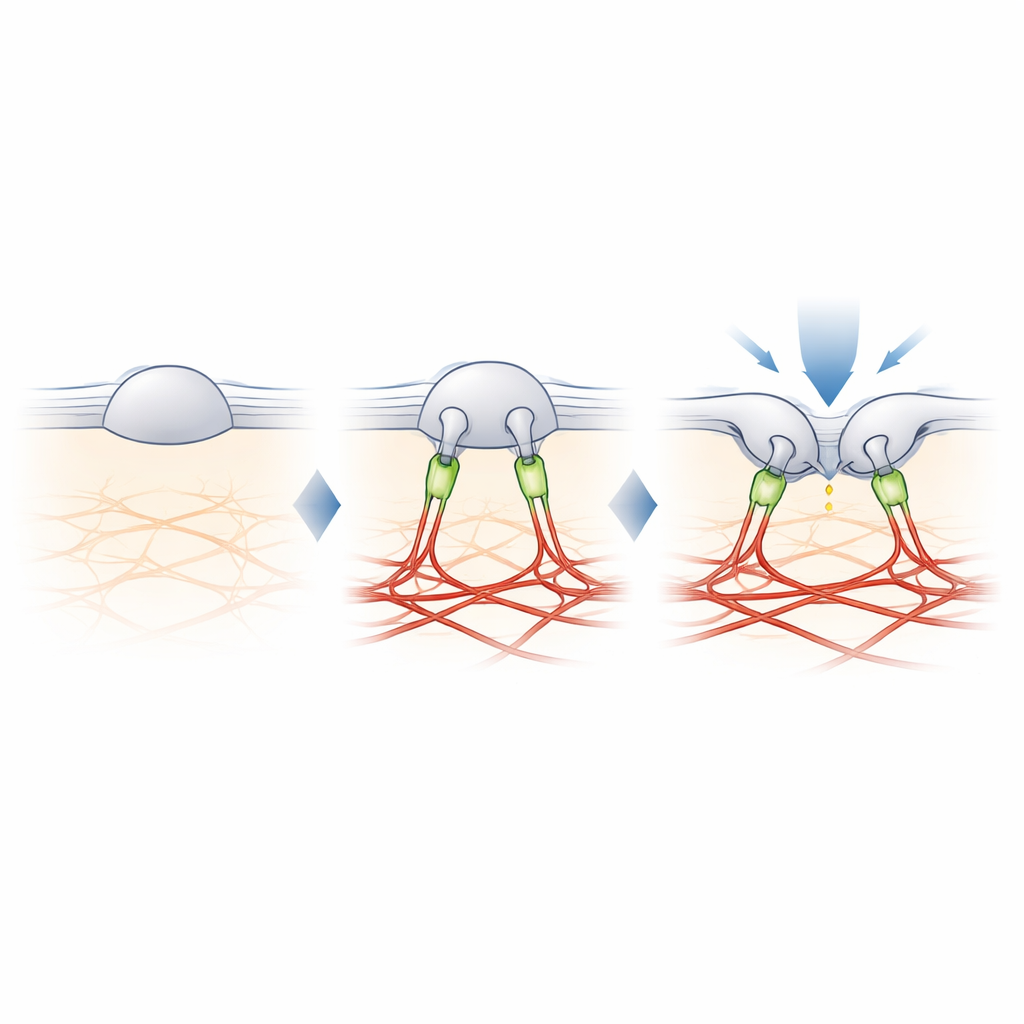
Filamin-B: The Tether that Tunes Touch
To find the missing link between PIEZO2 and actin, the authors cross-linked nearby proteins to PIEZO2 and identified them by mass spectrometry. Among several candidates, one stood out: filamin-B (FLNB), a large scaffold protein known to bind actin and many membrane proteins. Knocking down FLNB in engineered cells turned PIEZO2 into a robust stretch sensor and increased its mobility in the membrane, mimicking the effects of cutting its internal segment. Crucially, removing FLNB reduced PIEZO2’s sensitivity to poking with a blunt probe and raised the amount of indentation needed to open it. In sensory neurons from mice, partial loss of FLNB made the native PIEZO2 channels respond to membrane stretch they normally ignore, confirming that FLNB is essential for their usual preference for indentation.
From Molecular Tether to Sense of Touch
The study concludes that PIEZO2’s special role in touch comes from a physical tether to the actin scaffold, built in part from filamin-B. Instead of simply feeling how much the membrane is stretched, PIEZO2 feels how the membrane bends and shifts relative to this internal anchor, which keeps the channel slightly pre-opened and easier to trigger by a local poke. PIEZO1, by contrast, remains mostly untethered and responds mainly to overall tension. This molecular division of labor helps explain how our bodies use closely related channels to distinguish gentle touch from global forces such as blood flow or pressure. It also offers a blueprint for how cells in many tissues might fine-tune their mechanical sensitivity by adding or removing such tethers.
Citation: Mulhall, E.M., Yarishkin, O., Hill, R.Z. et al. The molecular basis of force selectivity by PIEZO2. Nature 653, 297–305 (2026). https://doi.org/10.1038/s41586-026-10182-7
Keywords: mechanosensation, PIEZO2, touch neurons, cytoskeleton, ion channels