Clear Sky Science · pl
Podstawy molekularne selektywności siły przez PIEZO2
Jak nasze nerwy rozróżniają pchnięcie od rozciągnięcia
Za każdym razem, gdy przesuwasz opuszkami palców po stronie lub czujesz, jak płuca rozszerzają się podczas oddechu, maleńkie maszyny molekularne w komórkach przekształcają siłę mechaniczną w sygnały elektryczne. Dwa blisko spokrewnione kanały, nazywane PIEZO1 i PIEZO2, odgrywają w tym procesie kluczową rolę. Reagują jednak na zupełnie różne rodzaje sił: jeden preferuje szerokie rozciąganie, drugi jest wyczulony na delikatne pchnięcia i naciski, które odbieramy jako dotyk. Artykuł ten ujawnia, jak PIEZO2 osiąga tę specjalizację, wskazując na fizyczne rusztowanie, które pomaga naszemu układowi nerwowemu rozróżniać rodzaje działających sił.
Dwoje rodzeństwa-czujników o różnych zadaniach
PIEZO1 i PIEZO2 to duże, przypominające trójnóg białka osadzone w błonie komórkowej, które otwierają się, gdy błona ulega odkształceniu. PIEZO1 występuje głównie w komórkach nie-neuronalnych, takich jak krwinki czerwone czy chrząstka, gdzie dobrze reaguje na ogólne rozciąganie lub przepływ płynów. PIEZO2, przeciwnie, skupia się w neuronach czucia dotyku i propriocepcji i jest niezbędny do działania delikatnego dotyku, orientacji ciała i oddychania. Pod mikroskopem ich struktury wyglądają zaskakująco podobnie, co utrudniało wyjaśnienie, dlaczego PIEZO1 reaguje silnie na rozciąganie błony, podczas gdy PIEZO2 znacznie lepiej odpowiada na lokalne wgłębienia, jak małe pchnięcie palcem po powierzchni komórki.
Obserwowanie ruchu pojedynczych cząsteczek pod działaniem siły
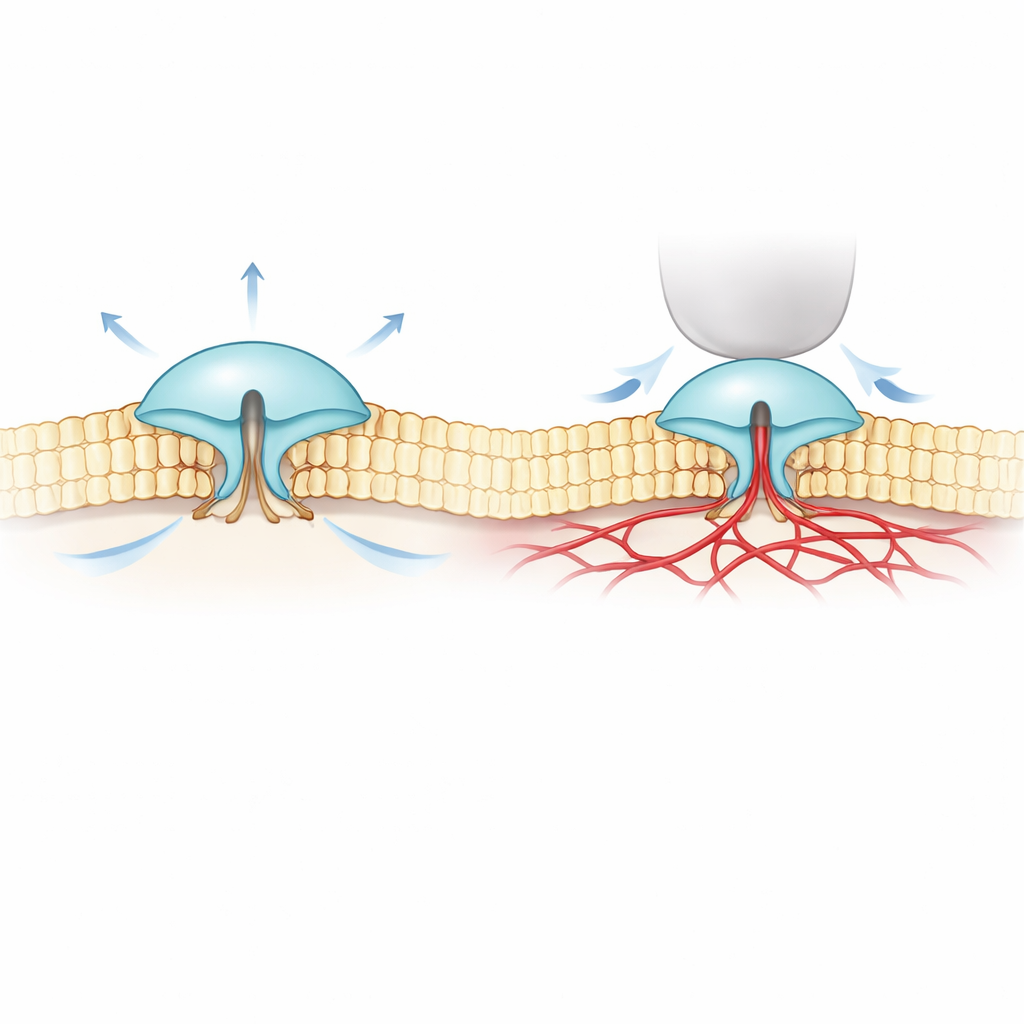
Aby rozwiązać tę zagadkę, autorzy połączyli ultradokładne obrazowanie fluorescencyjne (MINFLUX) z zapisem elektrycznym. Oznakowali wersje PIEZO1 i PIEZO2 niewielkimi światłoczułymi znacznikami na zewnętrznych częściach ich „łopatek” i obserwowali, jak zmienia się rozstaw tych łopatek w rzeczywistych błonach komórkowych. Stwierdzili, że PIEZO2 jest z natury bardziej zwarty i sztywniejszy niż PIEZO1: jego łopatki mniej fluktuują i spoczywają bliżej siebie w stanie spoczynku. Gdy komórki były pęczniejące, co zwiększało napięcie błony — podobnie jak nadmuchiwanie balonu — łopatki PIEZO1 rozsuwały się, a kanał otwierał się, pasując do prostej wizji „otwierania przez rozciąganie błony”. PIEZO2 zachowywał się odwrotnie. Jego łopatki lekko się zaciskały, a kanał praktycznie się nie otwierał, mimo że cała błona była napięta.
Ukryte połączenie z wewnętrznym szkieletem komórki
Te zaskakujące wyniki sugerowały, że PIEZO2 nie jest napędzany głównie przez otaczającą warstwę lipidową błony, lecz przez coś wewnątrz komórki. Zespół zwrócił uwagę na cytoszkielet aktynowy, sieć filamentów białkowych tuż pod błoną, która pomaga komórkom zachować kształt. W komórkach bogatych w aktynę zmniejszenie objętości komórki (co poluzowuje błonę i składa ją w siatkę aktynową) faktycznie powodowało rozszerzenie łopatek PIEZO2, znów w sposób przeciwny do reakcji PIEZO1. Gdy badacze chemicznie zakłócili aktynę lub usunęli elastyczny wewnętrzny fragment PIEZO2 wcześniej powiązany z dotykiem, zachowanie PIEZO2 odwróciło się: zaczął reagować na rozciąganie bardziej jak PIEZO1. Śledzenie pojedynczych cząsteczek wykazało ponadto, że normalny PIEZO2 praktycznie się nie przemieszcza w błonie — co zgadza się z jego unieruchomieniem — podczas gdy po usunięciu tego wewnętrznego fragmentu dyfunduje swobodnie.
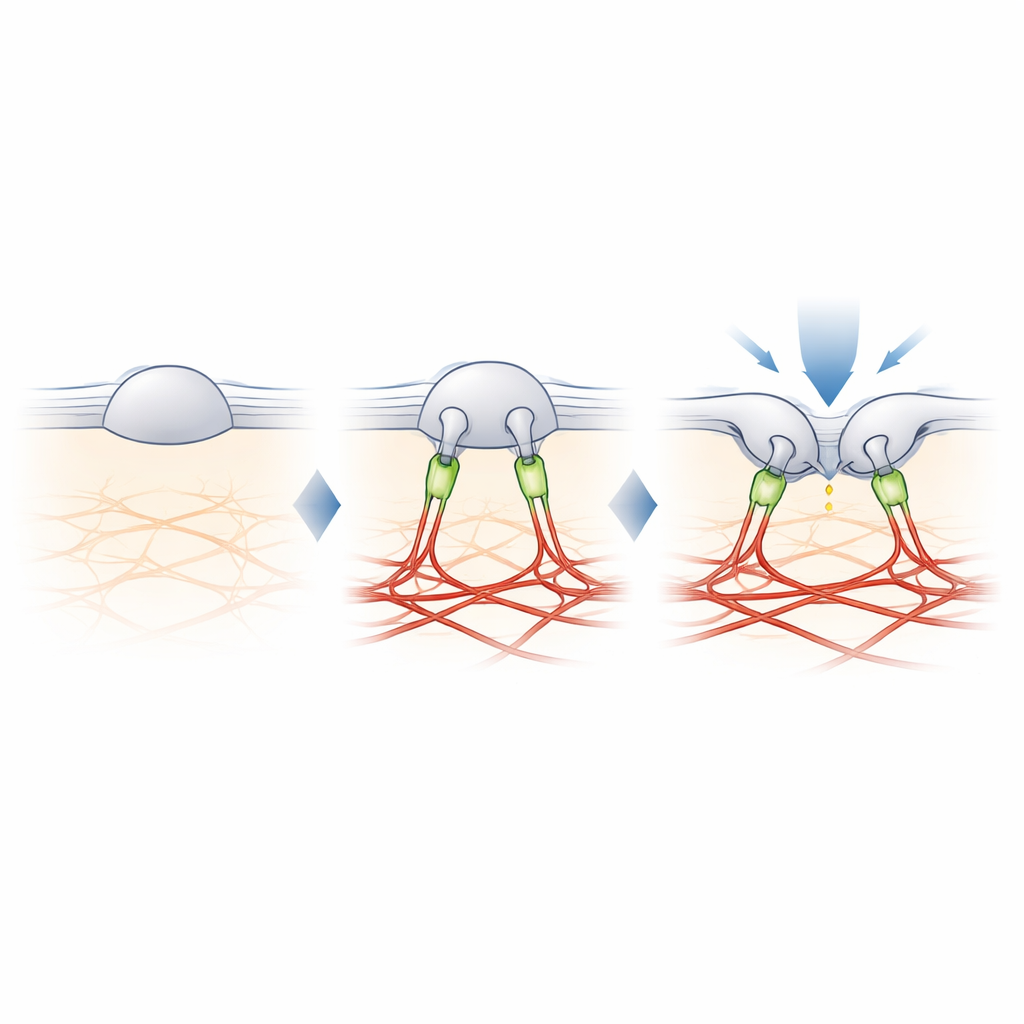
Filamina-B: tether, który stroi czucie dotyku
Aby znaleźć brakujące ogniwo między PIEZO2 a aktyną, autorzy skrzyżowali sąsiadujące białka z PIEZO2 i zidentyfikowali je za pomocą spektrometrii mas. Spośród kilku kandydatów wyróżniła się filamina-B (FLNB), duże białko-szkielet znane z wiązania aktyny i wielu białek błonowych. Zmniejszenie poziomu FLNB w komórkach inżynieryjnych przemieniło PIEZO2 w silny czujnik rozciągania i zwiększyło jego ruchliwość w błonie, naśladując efekty odcięcia wewnętrznego fragmentu. Co istotne, usunięcie FLNB zmniejszyło wrażliwość PIEZO2 na pchnięcia tęgim sondą i zwiększyło głębokość wgłębienia potrzebną do jego otwarcia. W neuronach czuciowych myszy częściowa utrata FLNB sprawiła, że natywne kanały PIEZO2 zaczęły reagować na rozciąganie błony, którego normalnie ignorują, co potwierdza, że FLNB jest niezbędna do ich zwykłej preferencji dla wgłębień.
Od molekularnego tethera do zmysłu dotyku
Badanie dochodzi do wniosku, że szczególna rola PIEZO2 w dotyku wynika z fizycznego przytwierdzenia do rusztowania aktynowego, w którym istotną rolę odgrywa filamina-B. Zamiast jedynie odczuwać, jak bardzo błona jest rozciągnięta, PIEZO2 wykrywa, jak błona zgina się i przesuwa względem tego wewnętrznego kotwiczenia, co utrzymuje kanał lekko wstępnie otwartym i ułatwia jego wyzwalanie przez lokalne pchnięcie. PIEZO1 natomiast pozostaje w dużej mierze nieprzytwierdzony i reaguje głównie na ogólne napięcie. Ten molekularny podział pracy pomaga wyjaśnić, w jaki sposób organizm używa blisko spokrewnionych kanałów do rozróżniania delikatnego dotyku od globalnych sił, takich jak przepływ krwi czy ciśnienie. Otwiera to także drogę do zrozumienia, jak komórki w różnych tkankach mogą precyzyjnie dostroić swoją czułość mechaniczną przez dodawanie lub usuwanie takich tetherów.
Cytowanie: Mulhall, E.M., Yarishkin, O., Hill, R.Z. et al. The molecular basis of force selectivity by PIEZO2. Nature 653, 297–305 (2026). https://doi.org/10.1038/s41586-026-10182-7
Słowa kluczowe: mechanosensacja, PIEZO2, neurony dotykowe, cytoszkielet, kanały jonowe