Clear Sky Science · pt
A base molecular da seletividade de força pelo PIEZO2
Como nossos nervos distinguem um toque de um estiramento
Cada vez que você desliza as pontas dos dedos sobre uma página ou sente seus pulmões se expandirem ao respirar, pequenas máquinas moleculares nas células convertem força mecânica em sinais elétricos. Dois canais intimamente relacionados, chamados PIEZO1 e PIEZO2, são centrais nesse processo. Ainda assim, eles respondem a tipos de forças muito diferentes: um prefere um estiramento amplo, o outro é ajustado a pokes e pressões suaves que percebemos como toque. Este artigo revela como o PIEZO2 alcança essa especialização, mostrando um cabo físico que ajuda nosso sistema nervoso a distinguir um tipo de força de outro.
Dois sensores irmãos com funções distintas
PIEZO1 e PIEZO2 são grandes proteínas em formato de tripé que se situam na membrana celular e se abrem quando a membrana é deformada. PIEZO1 é encontrado principalmente em células não nervosas, como hemácias e cartilagem, onde responde bem a estiramento geral ou fluxo de líquido. PIEZO2, em contraste, está concentrado em neurônios sensoriais de toque e propriocepção e é crucial para o tato suave, a consciência corporal e a respiração. Ao microscópio, suas estruturas parecem notavelmente semelhantes, o que torna intrigante o fato de o PIEZO1 reagir fortemente ao estiramento da membrana, enquanto o PIEZO2 responde muito melhor à indentação local, como um pequeno toque na superfície celular.
Observando moléculas individuais se moverem sob força
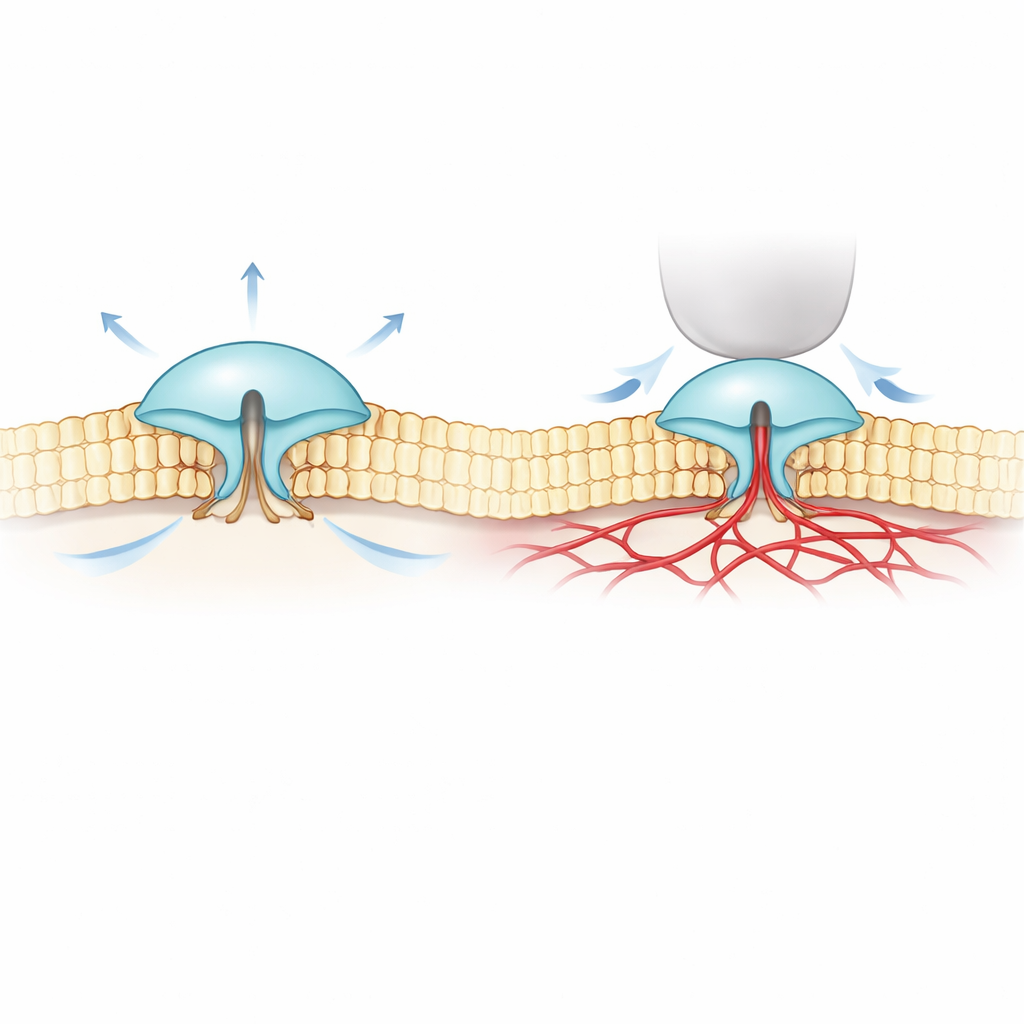
Para abordar esse mistério, os autores combinaram imagem fluorescente ultraprécisa (chamada MINFLUX) com registros elétricos. Eles projetaram versões do PIEZO1 e PIEZO2 que carregam pequenas marcas sensíveis à luz nas partes externas de suas “lâminas” e observaram como o espaçamento dessas lâminas mudava nas membranas celulares reais. Descobriram que o PIEZO2 é intrinsecamente mais compacto e rígido que o PIEZO1: suas lâminas flutuam menos e ficam mais próximas em repouso. Quando as células foram inchadas para aumentar a tensão da membrana — semelhante a inflar um balão — as lâminas do PIEZO1 se espalharam e o canal abriu, confirmando a ideia simples de que ele é “puxado aberto pela membrana”. O PIEZO2 se comportou de forma oposta. Suas lâminas apertaram-se ligeiramente e o canal mal abriu, embora toda a membrana estivesse sob tensão.
Uma conexão oculta com o arcabouço interno da célula
Esses resultados surpreendentes sugeriram que o PIEZO2 não é acionado principalmente pela camada gordurosa circundante da membrana, mas por algo dentro da célula. A equipe voltou-se para o citoesqueleto de actina, uma malha de filamentos proteicos logo abaixo da membrana que ajuda a dar forma às células. Em um tipo celular rico em actina, reduzir o volume celular (o que frouxeia a membrana e a dobra na malha de actina) na verdade fez com que as lâminas do PIEZO2 se expandissem, novamente em oposição ao PIEZO1. Quando os pesquisadores desorganizaram quimicamente a actina, ou deletaram um segmento interno flexível do PIEZO2 anteriormente ligado ao tato, o comportamento do PIEZO2 virou: ele passou a responder ao estiramento muito mais parecido com o PIEZO1. Rastreamento de moléculas únicas mostrou ainda que o PIEZO2 normal praticamente não se move na membrana — consistente com estar ancorado — enquanto após a remoção desse segmento interno ele difunde-se livremente.
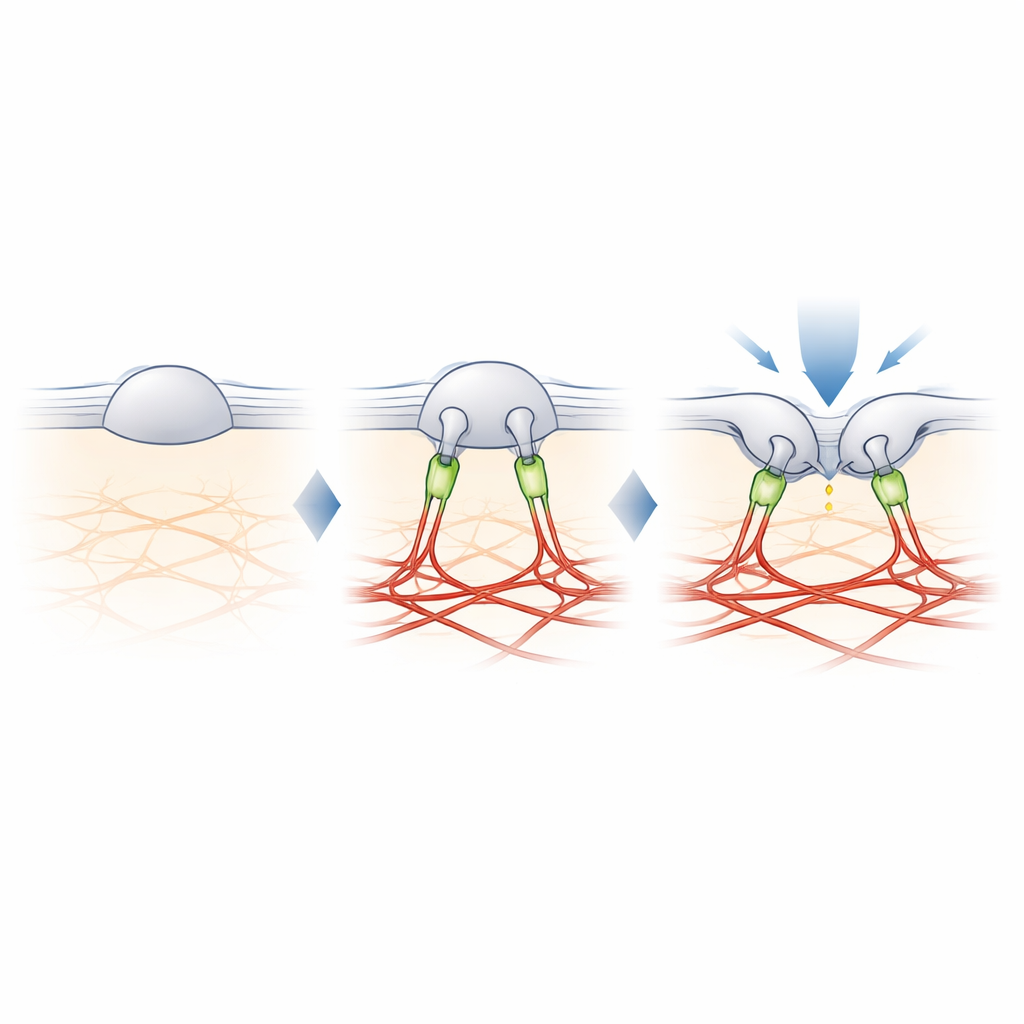
Filamina-B: o cabo que ajusta o tato
Para encontrar o elo perdido entre o PIEZO2 e a actina, os autores entrecruzaram proteínas próximas ao PIEZO2 e as identificaram por espectrometria de massas. Entre vários candidatos, um se destacou: filamina-B (FLNB), uma grande proteína de andaime conhecida por se ligar à actina e a muitas proteínas de membrana. Reduzir os níveis de FLNB em células modificadas transformou o PIEZO2 em um sensor robusto de estiramento e aumentou sua mobilidade na membrana, mimetizando os efeitos de cortar seu segmento interno. Crucialmente, remover a FLNB diminuiu a sensibilidade do PIEZO2 a toques com uma sonda romba e aumentou a quantidade de indentação necessária para abri-lo. Em neurônios sensoriais de camundongos, a perda parcial de FLNB fez com que os canais PIEZO2 nativos respondessem ao estiramento da membrana que normalmente ignoram, confirmando que a FLNB é essencial para a preferência usual por indentação.
Do cabo molecular ao sentido do tato
O estudo conclui que o papel especial do PIEZO2 no tato deriva de um cabo físico ao arcabouço de actina, composto em parte pela filamina-B. Em vez de simplesmente sentir quanto a membrana é esticada, o PIEZO2 percebe como a membrana dobra e se desloca em relação a essa âncora interna, que mantém o canal ligeiramente pré-aberto e mais fácil de acionar por um toque local. O PIEZO1, em contraste, permanece em grande parte sem ancoragem e responde principalmente à tensão geral. Essa divisão molecular de tarefas ajuda a explicar como nossos corpos usam canais intimamente relacionados para distinguir o toque suave de forças globais, como fluxo sanguíneo ou pressão. Também oferece um roteiro sobre como células em muitos tecidos podem ajustar sua sensibilidade mecânica ao adicionar ou remover esses cabos.
Citação: Mulhall, E.M., Yarishkin, O., Hill, R.Z. et al. The molecular basis of force selectivity by PIEZO2. Nature 653, 297–305 (2026). https://doi.org/10.1038/s41586-026-10182-7
Palavras-chave: mecanossensação, PIEZO2, neurônios do tato, citoesqueleto, canais iônicos