Clear Sky Science · nl
De moleculaire basis van krachtspecificiteit door PIEZO2
Hoe onze zenuwen een prik van een rek onderscheiden
Elke keer dat je vingertoppen over een pagina strijken of je longen uitzetten tijdens het ademen, zetten kleine moleculaire machines in je cellen mechanische kracht om in elektrische signalen. Twee nauw verwante kanalen, PIEZO1 en PIEZO2 genoemd, staan centraal in dit proces. Toch reageren ze op heel verschillende soorten krachten: de ene geeft de voorkeur aan brede rek, de andere is afgestemd op zachte prikken en drukken die wij als aanraking ervaren. Dit artikel onthult hoe PIEZO2 die specialisatie bereikt en toont een fysieke houvast die ons zenuwstelsel helpt het ene type kracht van het andere te onderscheiden.
Twee verwante sensoren met verschillende taken
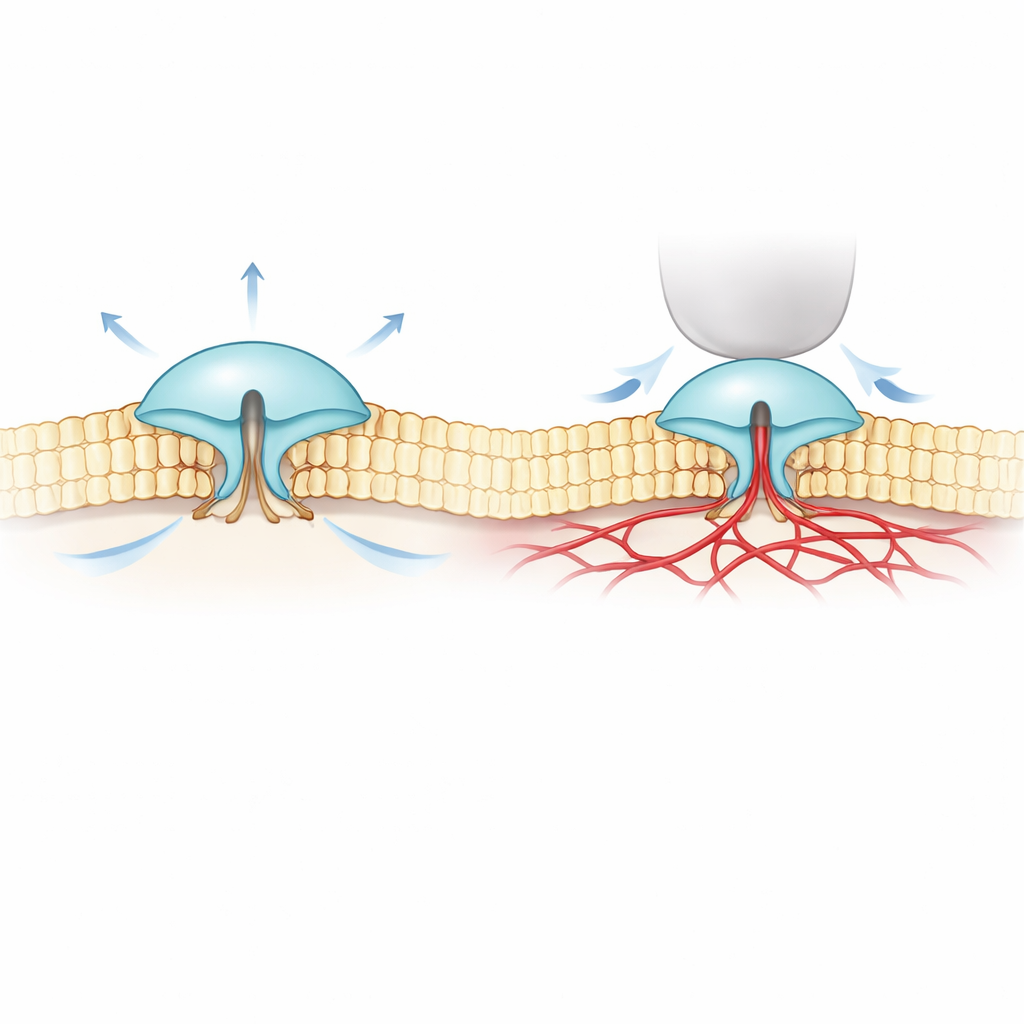
PIEZO1 en PIEZO2 zijn grote, driepootachtige eiwitten die in het celmembraan zitten en openen wanneer het membraan vervormd wordt. PIEZO1 komt vooral voor in niet-zenuwachtige cellen zoals rode bloedcellen en kraakbeen, waar het goed reageert op algemene rek of stroming van vloeistof. PIEZO2 daarentegen concentreert zich in aanraking- en houdingszintuigcellen en is cruciaal voor zachte aanraking, lichaamsbewustzijn en ademhaling. Onder de microscoop lijken hun structuren opvallend veel op elkaar, wat het raadselachtig maakte waarom PIEZO1 sterk reageert op membraanrek, terwijl PIEZO2 veel beter reageert op lokale inkeping, zoals een klein vingertje dat in het celoppervlak prikt.
Enkele moleculen zien bewegen onder kracht
Om dit mysterie te ontrafelen combineerden de auteurs extreem precieze fluorescentie-imaging (MINFLUX) met elektrische opnames. Ze maakten versies van PIEZO1 en PIEZO2 met kleine lichtgevoelige labels op de buitenste delen van hun “bladen” en observeerden hoe de afstand tussen die bladen veranderde in echte celmembranen. Ze vonden dat PIEZO2 van zichzelf compacter en stijver is dan PIEZO1: zijn bladen fluctueren minder en liggen in rust dichter bij elkaar. Toen cellen opgezwollen werden om de membraanspanning te verhogen—vergelijkbaar met het opblazen van een ballon—spreidden de bladen van PIEZO1 uit en het kanaal opende, wat past bij het simpele beeld van ‘door het membraan opengetrokken’. PIEZO2 gedroeg zich juist tegengesteld. Zijn bladen trokken iets steviger samen en het kanaal opende nauwelijks, ook al stond het hele membraan onder spanning.
Een verborgen verbinding met het binnenste steigerwerk van de cel
Deze verrassende resultaten suggereerden dat PIEZO2 niet voornamelijk wordt aangedreven door de omringende vetlaag van het membraan, maar door iets anders binnenin de cel. Het team richtte zich op het actine-cytoskelet, een netwerk van eiwitfilamenten direct onder het membraan dat helpt de vorm van cellen te behouden. In een celtype rijk aan actine zorgde het verkleinen van het celvolume (waardoor het membraan slapper wordt en in het actinenet wordt gevouwen) er juist voor dat de bladen van PIEZO2 uitzetten, weer het tegenovergestelde van PIEZO1. Wanneer de onderzoekers actine chemisch verstoorden, of een labiele interne segment van PIEZO2 verwijderden dat eerder aan aanraking was gekoppeld, keerde het gedrag van PIEZO2 om: nu begon het veel meer op PIEZO1 te reageren bij rek. Single-molecule tracking toonde verder aan dat normaal PIEZO2 nauwelijks beweegt in het membraan—consistent met verankering—terwijl het na verstoring van dit interne segment vrij diffundeert.
Filamine-B: de tether die aanraking afstemt
Om de ontbrekende schakel tussen PIEZO2 en actine te vinden, kruisverbonden de auteurs nabijgelegen eiwitten aan PIEZO2 en identificeerden deze met massaspectrometrie. Onder verschillende kandidaten stak één er met kop en schouders bovenuit: filamine-B (FLNB), een groot scaffold-eiwit dat bekendstaat om het binden van actine en veel membraaneiwitten. Het onderdrukken van FLNB in gemodificeerde cellen veranderde PIEZO2 in een robuuste rek-sensor en verhoogde zijn beweeglijkheid in het membraan, wat de effecten nabootste van het verwijderen van het interne segment. Cruciaal was dat het wegnemen van FLNB de gevoeligheid van PIEZO2 voor het prikken met een stomp werktuigje verminderde en de noodzakelijke inkeping om het te openen vergrootte. In sensorische neuronen van muizen maakte gedeeltelijk verlies van FLNB dat de native PIEZO2-kanalen gingen reageren op membraanrek dat ze normaal negeren, waarmee werd bevestigd dat FLNB essentieel is voor hun gebruikelijke voorkeur voor inkeping.
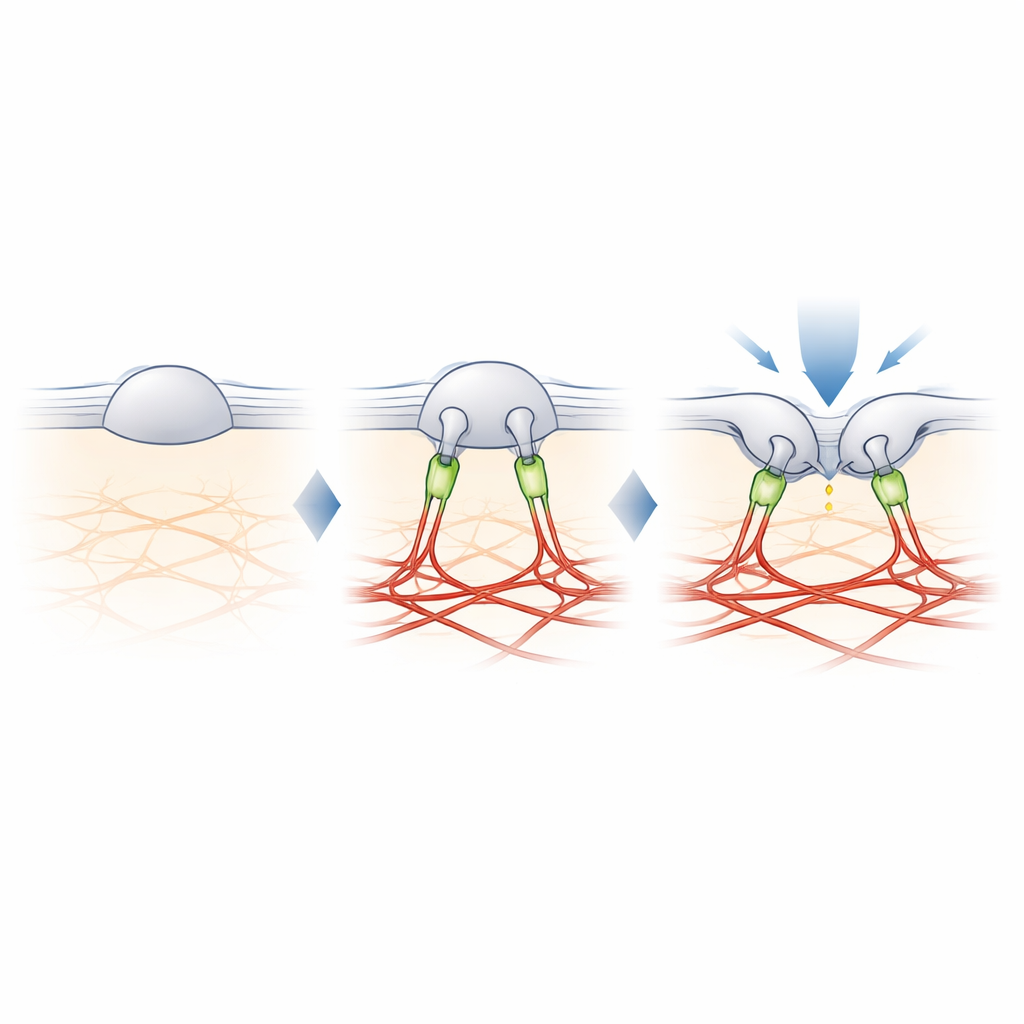
Van moleculaire tether naar gevoel van aanraking
De studie concludeert dat PIEZO2’s bijzondere rol in aanraking voortkomt uit een fysieke tether aan het actine-steigerwerk, deels opgebouwd uit filamine-B. In plaats van alleen te voelen hoeveel het membraan wordt uitgerekt, voelt PIEZO2 hoe het membraan buigt en verschuift ten opzichte van dit interne anker, wat het kanaal licht voorgeopend houdt en gemakkelijker te activeren maakt door een lokale prik. PIEZO1 daarentegen blijft grotendeels niet-verankerd en reageert vooral op algemene spanning. Deze moleculaire arbeidsdeling helpt verklaren hoe ons lichaam nauw verwante kanalen gebruikt om zachte aanraking te onderscheiden van globale krachten zoals bloedstroom of druk. Het biedt ook een blauwdruk voor hoe cellen in vele weefsels hun mechanische gevoeligheid kunnen bijstellen door zulke tethers toe te voegen of te verwijderen.
Bronvermelding: Mulhall, E.M., Yarishkin, O., Hill, R.Z. et al. The molecular basis of force selectivity by PIEZO2. Nature 653, 297–305 (2026). https://doi.org/10.1038/s41586-026-10182-7
Trefwoorden: mechanosensatie, PIEZO2, toucheuronen, cytoskelet, ionkanalen