Clear Sky Science · fr
La base moléculaire de la sélectivité de force par PIEZO2
Comment nos nerfs distinguent une piqûre d'un étirement
Chaque fois que vous frottez le bout des doigts sur une page ou sentez vos poumons se dilater en respirant, de minuscules machines moléculaires dans vos cellules convertissent la force mécanique en signaux électriques. Deux canaux étroitement apparentés, appelés PIEZO1 et PIEZO2, sont au cœur de ce processus. Pourtant, ils répondent à des types de forces très différents : l’un préfère les étirements larges, l’autre est accordé aux pressions et picotements légers que nous percevons comme le toucher. Cet article révèle comment PIEZO2 réalise cette spécialisation, en dévoilant un lien physique qui aide notre système nerveux à distinguer un type de force d’un autre.
Deux capteurs « frères » aux fonctions distinctes
PIEZO1 et PIEZO2 sont de grandes protéines en forme de trépied qui s’installent dans la membrane cellulaire et s’ouvrent lorsque la membrane est déformée. PIEZO1 se trouve principalement dans des cellules non nerveuses comme les globules rouges et le cartilage, où il répond bien à l’étirement global ou au flux de fluide. PIEZO2, en revanche, est concentré dans les neurones du toucher et de la proprioception et est crucial pour le toucher fin, la conscience corporelle et la respiration. Au microscope, leurs structures paraissent étonnamment similaires, ce qui a rendu énigmatique le fait que PIEZO1 réagisse fortement à l’étirement membranaire, alors que PIEZO2 réponde bien mieux à une indentation locale, comme une petite piqûre sur la surface cellulaire.
Observer des molécules individuelles sous la force
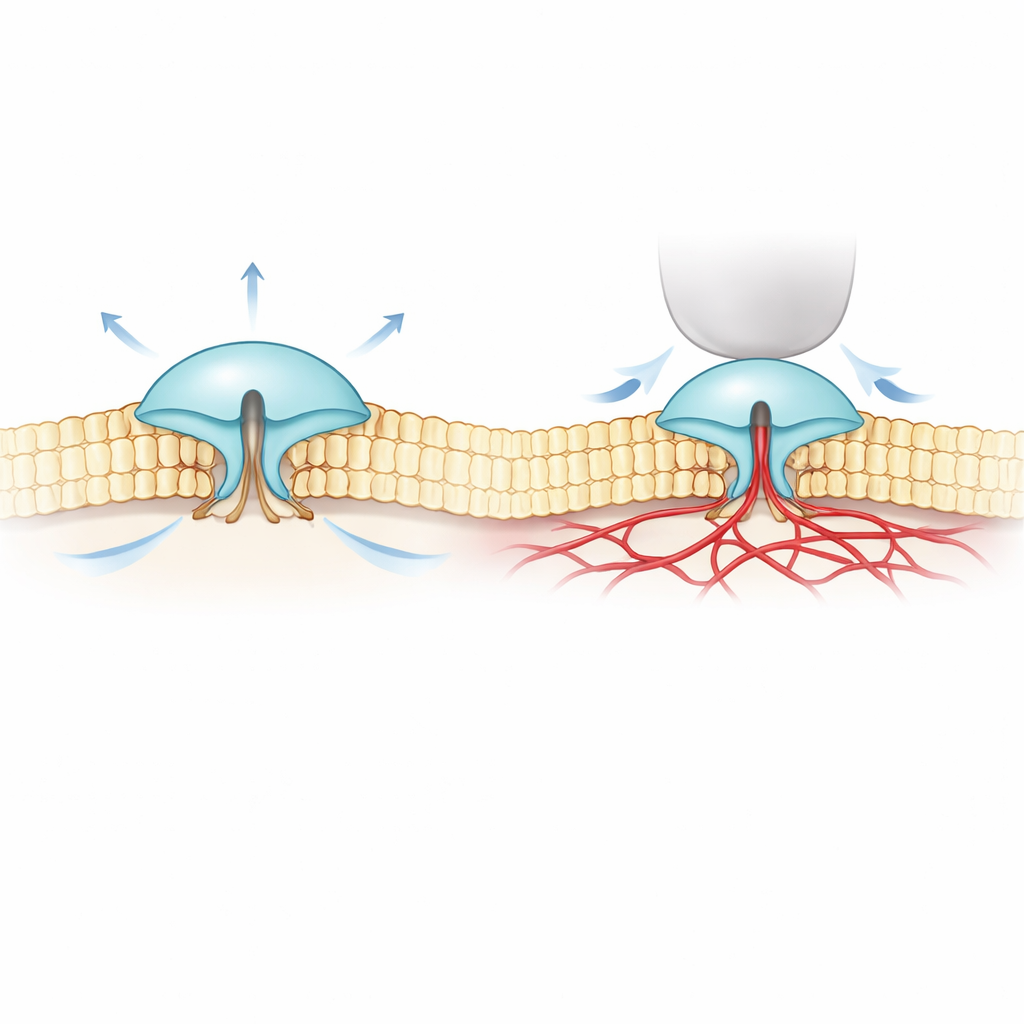
Pour résoudre ce mystère, les auteurs ont combiné une imagerie fluorescente ultra-précise (appelée MINFLUX) à des enregistrements électriques. Ils ont conçu des versions de PIEZO1 et PIEZO2 portant de petites étiquettes sensibles à la lumière sur les parties externes de leurs « lames » et ont observé comment l’écartement de ces lames changeait dans de vraies membranes cellulaires. Ils ont constaté que PIEZO2 est intrinsèquement plus compact et rigide que PIEZO1 : ses lames fluctuent moins et restent plus rapprochées au repos. Lorsque les cellules ont été gonflées pour augmenter la tension membranaire — un peu comme gonfler un ballon — les lames de PIEZO1 se sont écartées et le canal s’est ouvert, conforme à l’idée simple d’un canal « tiré » par la membrane. PIEZO2 s’est comporté de façon opposée. Ses lames se sont légèrement resserrées et le canal s’est à peine ouvert, même si toute la membrane était sous tension.
Un lien caché avec l’armature interne de la cellule
Ces résultats surprenants suggéraient que PIEZO2 n’est pas principalement actionné par la couche lipidique environnante de la membrane mais par quelque chose d’autre à l’intérieur de la cellule. L’équipe a regardé du côté du cytosquelette d’actine, un réseau de filaments protéiques juste sous la membrane qui aide à donner sa forme à la cellule. Dans un type cellulaire riche en actine, réduire le volume cellulaire (ce qui relâche la membrane et la replie dans le maillage d’actine) a en fait provoqué l’expansion des lames de PIEZO2, encore une fois à l’opposé de PIEZO1. Lorsque les chercheurs ont perturbé chimiquement l’actine, ou supprimé un segment interne flexible de PIEZO2 auparavant lié au toucher, le comportement de PIEZO2 a basculé : il a alors commencé à répondre à l’étirement de façon beaucoup plus semblable à PIEZO1. Le suivi de molécules individuelles a montré en outre que le PIEZO2 normal se déplace à peine dans la membrane — cohérent avec une ancrage — alors qu’après suppression de ce segment interne il diffuse librement.
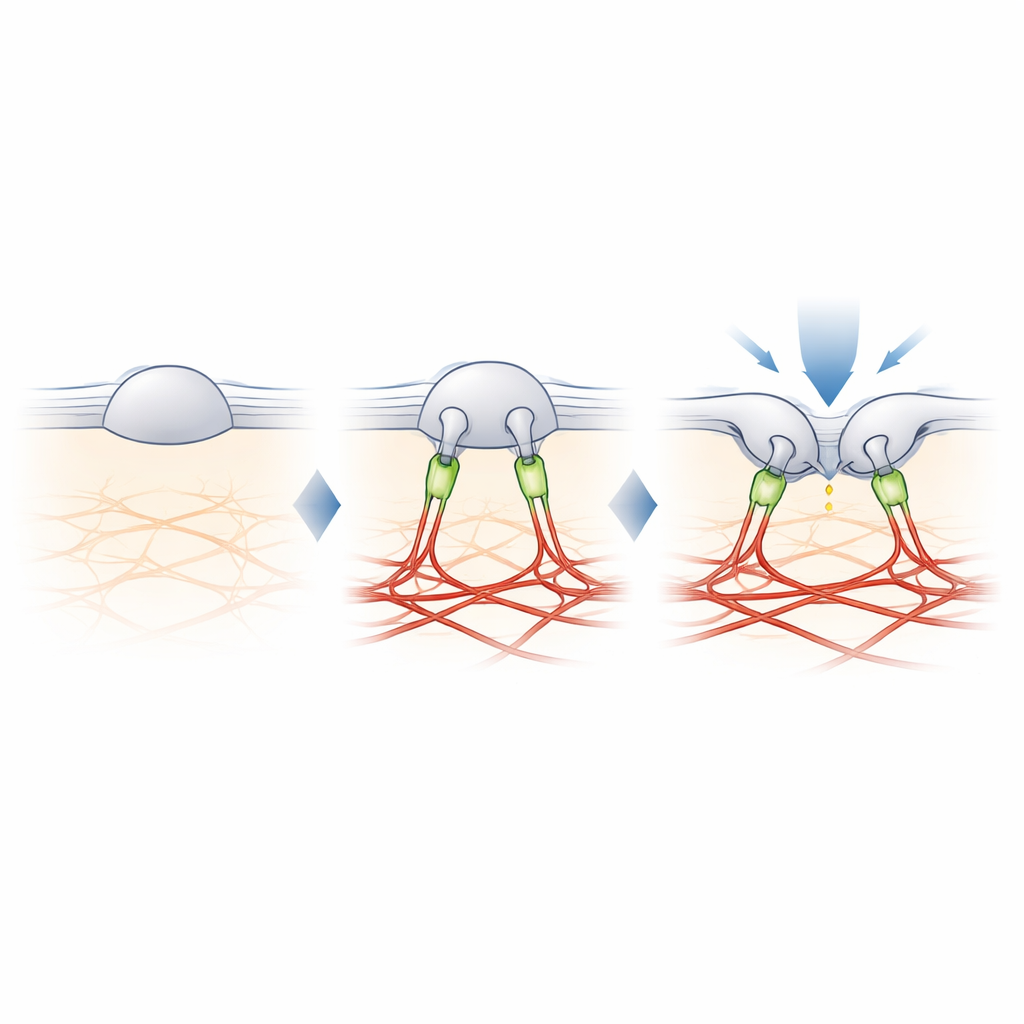
Filamine-B : l’attache qui règle le toucher
Pour trouver le lien manquant entre PIEZO2 et l’actine, les auteurs ont rétrocroisé les protéines proches de PIEZO2 puis les ont identifiées par spectrométrie de masse. Parmi plusieurs candidats, une protéine a retenu l’attention : la filamine-B (FLNB), une grosse protéine échafaudage connue pour lier l’actine et de nombreuses protéines membranaires. Réduire l’expression de FLNB dans des cellules modifiées a transformé PIEZO2 en un détecteur d’étirement robuste et a accru sa mobilité dans la membrane, mimant les effets de la coupure de son segment interne. De façon cruciale, l’élimination de FLNB a réduiit la sensibilité de PIEZO2 aux picotements avec une sonde émoussée et augmenté la profondeur d’indentation nécessaire pour l’ouvrir. Dans des neurones sensoriels de souris, une perte partielle de FLNB a rendu les canaux PIEZO2 natifs réactifs à l’étirement membranaire qu’ils ignoraient habituellement, confirmant que FLNB est essentiel à leur préférence habituelle pour l’indentation.
Du lien moléculaire au sens du toucher
L’étude conclut que le rôle particulier de PIEZO2 dans le toucher provient d’un ancrage physique au réseau d’actine, en partie assuré par la filamine-B. Plutôt que de simplement mesurer l’étirement global de la membrane, PIEZO2 détecte la façon dont la membrane se plie et se déplace par rapport à cette ancre interne, ce qui maintient le canal légèrement pré-ouvert et plus facile à déclencher par une pression locale. PIEZO1, en revanche, reste majoritairement non ancré et répond principalement à la tension globale. Cette division moléculaire du travail aide à expliquer comment notre organisme utilise des canaux proches pour distinguer le toucher léger des forces globales telles que le flux sanguin ou la pression. Elle offre aussi une feuille de route pour comprendre comment les cellules de nombreux tissus pourraient ajuster leur sensibilité mécanique en ajoutant ou retirant de tels ancrages.
Citation: Mulhall, E.M., Yarishkin, O., Hill, R.Z. et al. The molecular basis of force selectivity by PIEZO2. Nature 653, 297–305 (2026). https://doi.org/10.1038/s41586-026-10182-7
Mots-clés: mécanosensation, PIEZO2, neurones du toucher, cytosquelette, canaux ioniques