Clear Sky Science · pt
Estruturas pós-catálise do complexo I mitocondrial com ubiquinol-10 ligado ao sítio ativo
Como Nossas Células Transformam Alimento em Combustível
As mitocôndrias são frequentemente chamadas de usinas de energia das nossas células, e uma de suas máquinas-chave, conhecida como complexo I, inicia o processo de converter a energia dos alimentos na molécula ATP. Este estudo foca o complexo I em detalhe atômico para ver exatamente como ele se apresenta logo após realizar seu trabalho, quando uma pequena molécula gordurosa chamada ubiquinona-10 acabou de ser convertida em sua forma carregada e rica em energia. Entender esse momento fugaz ajuda a explicar como nossas células capturam energia com tanta eficiência e por que falhas nessa máquina podem contribuir para doenças do cérebro, músculos e coração.
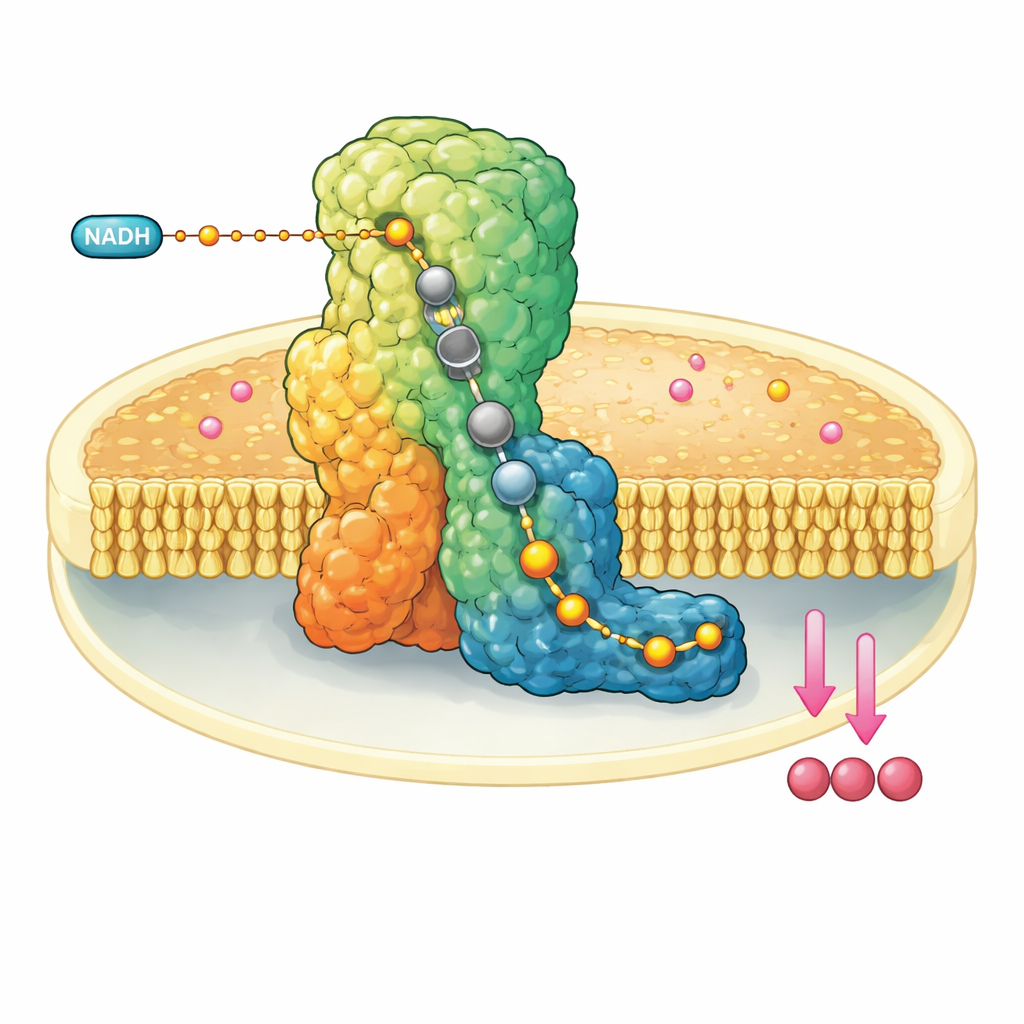
Uma Máquina Molecular na Parede Mitocondrial
O complexo I é um enorme conjunto proteico em forma de L embutido na membrana interna das mitocôndrias. Ele retira elétrons de uma molécula combustível, o NADH, e os transfere por um fio interno de clusters metálicos até alcançarem a ubiquinona-10, um transportador móvel que leva os elétrons adiante na cadeia respiratória. Ao mesmo tempo, o complexo I usa a energia liberada para mover quatro prótons através da membrana, ajudando a construir um gradiente eletroquímico que, em última instância, alimenta a síntese de ATP. Por causa desse papel central, o mau funcionamento do complexo I está ligado a uma série de distúrbios neuromusculares e metabólicos e a danos teciduais durante interrupções no suprimento sanguíneo.
Congelando a Máquina no Meio de um Ciclo
Os pesquisadores reconstituíram o complexo I bovino em pequenos discos de membrana artificial que também continham ubiquinona-10. Em seguida, misturaram rapidamente um grande excesso de NADH e mergulharam a amostra em nitrogênio líquido em questão de dezenas de segundos, aprisionando a enzima após ela ter realizado pelo menos um ciclo catalítico. Usando microscopia crioeletrônica de última geração com resoluções de até cerca de dois angströms—suficiente para visualizar moléculas individuais de água—foi possível reconstruir múltiplas estruturas tridimensionais do complexo I em diferentes estados de repouso. Crucialmente, na forma fechada e ativa, observaram ubiquinol-10, o produto reduzido, ocupando completamente todo o comprimento do estreito canal onde a reação ocorre.
Seguindo o Caminho do Transportador de Energia
Na forma ativa, a equipe identificou duas conformações distintas da mesma molécula de ubiquinol-10 acomodada no final de seu canal de ligação, perto do cluster terminal ferro-enxofre. Comparado a estruturas anteriores com ubiquinona oxidada, a cabeça da molécula deslocou posição e orientação, formando uma forte ligação de hidrogênio com uma cadeia lateral de tirosina específica, mas ficando longe demais de uma histidina próxima para interagir diretamente. Simulações computacionais complementares das cargas circundantes e das moléculas de água mostraram que essa configuração é mais estável quando tanto essa tirosina quanto um resíduo de aspartato estão desprotonados—coerente com o fato de terem acabado de doar os dois prótons que convertem ubiquinona em ubiquinol. A segunda posição, definida de forma mais fraca, provavelmente representa uma média de vários estados produtos intimamente relacionados enquanto o ubiquinol se prepara para deixar o sítio.
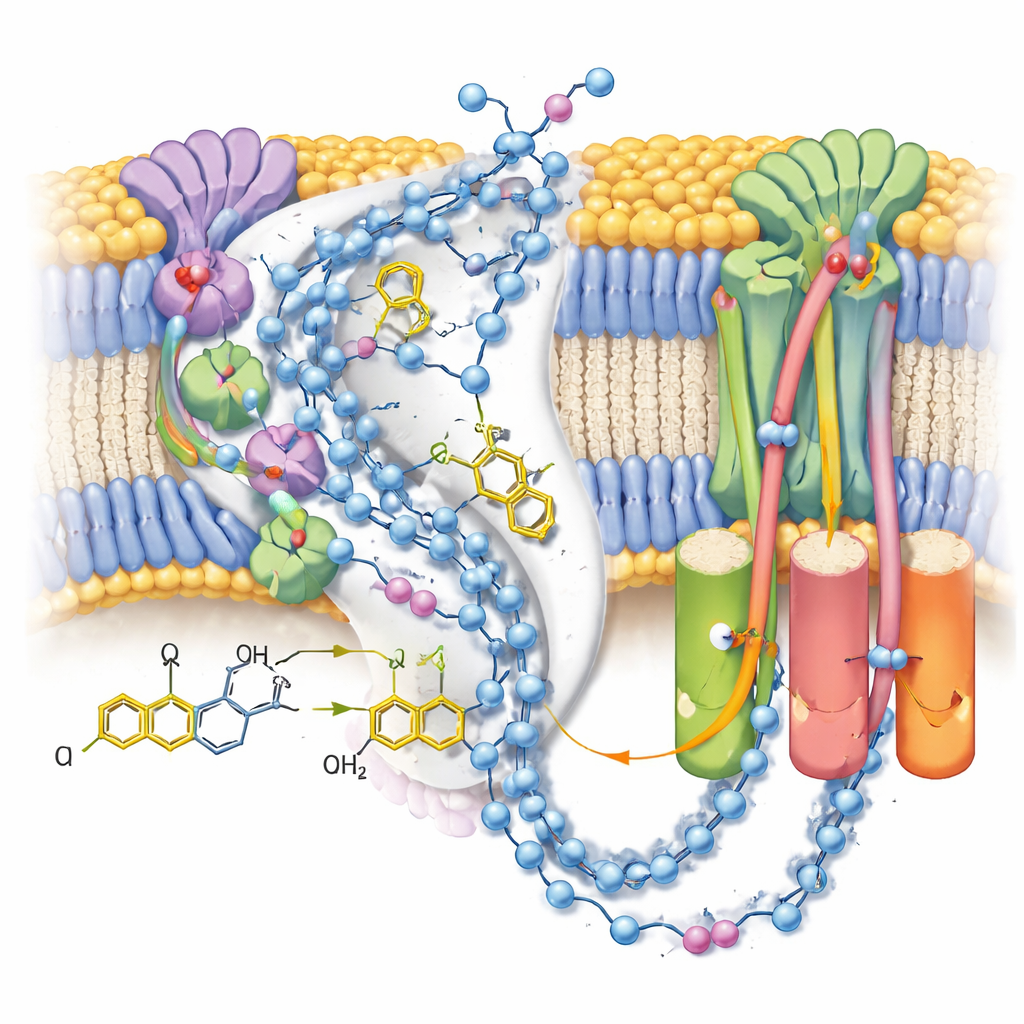
Mapeando as Vias de Prótons
Como os mapas revelaram muitas moléculas de água ordenadas, os autores puderam traçar cadeias quase contínuas de resíduos polares e água através do braço da membrana do complexo I. Essas cadeias conectam o sítio de reação da ubiquinona a três módulos separados de tipo “antiportador” que transportam prótons pela membrana. No estado fechado pós-reação com ubiquinol ligado, as conexões parecem em grande parte intactas, mas pontuadas por pequenas lacunas onde a água provavelmente é demasiado móvel para ser vista, sugerindo caminhos flexíveis em vez de tubos rígidos. Em contraste, um estado alternativo aberto e desativado—estabilizado aqui por uma molécula detergente alojada no canal—mostra uma hélice dobrada que interrompe uma conexão chave, apoiando a ideia de que esse estado regulatório bloqueia temporariamente a transferência de prótons em longa distância.
O Que Isso Significa para a Saúde Celular
No conjunto, as imagens estruturais e as simulações sustentam uma sequência na qual a ubiquinona se liga em uma pose pré-reação, aceita dois elétrons e dois prótons para tornar-se ubiquinol, e então permanece em uma pose pós-reação enquanto os grupos proteicos próximos aguardam ser reprotonados pelo lado da matriz da membrana. O trabalho esclarece como mudanças sutis na posição e na carga de alguns átomos no sítio ativo podem repercutir por longos caminhos de prótons para acionar o bombeamento, e como formas alternativas, desativadas, podem interromper esse fluxo. Ao aprimorar nossa visão do complexo I em ação, esses resultados oferecem um quadro para entender defeitos herdados, a ação de fármacos e o dano mitocondrial que ocorre durante eventos como ataques cardíacos e AVCs.
Citação: Chung, I., Pereira, C.S., Wright, J.J. et al. Post-catalysis structures of mitochondrial complex I with ubiquinol-10 bound in the active site. Nat Commun 17, 3506 (2026). https://doi.org/10.1038/s41467-026-70030-0
Palavras-chave: complexo I mitocondrial, ubiquinona-10, estrutura por crio-ME, bombeamento de prótons, fosforilação oxidativa