Clear Sky Science · fr
Structures post‑catalytiques du complexe I mitochondrial avec l’ubiquinol‑10 lié au site actif
Comment nos cellules transforment la nourriture en énergie
Les mitochondries sont souvent décrites comme les centrales électriques de nos cellules, et l’une de leurs machines clés, appelée complexe I, initie la conversion de l’énergie contenue dans les aliments en la molécule ATP. Cette étude examine le complexe I au niveau atomique pour voir exactement à quoi il ressemble juste après avoir accompli son travail, lorsqu’une petite molécule lipophile appelée ubiquinone‑10 vient d’être convertie en sa forme chargée et riche en énergie. Comprendre cet instant éphémère aide à expliquer comment nos cellules captent l’énergie de façon si efficace et pourquoi les perturbations de cette machinerie peuvent contribuer à des maladies du cerveau, des muscles et du cœur.
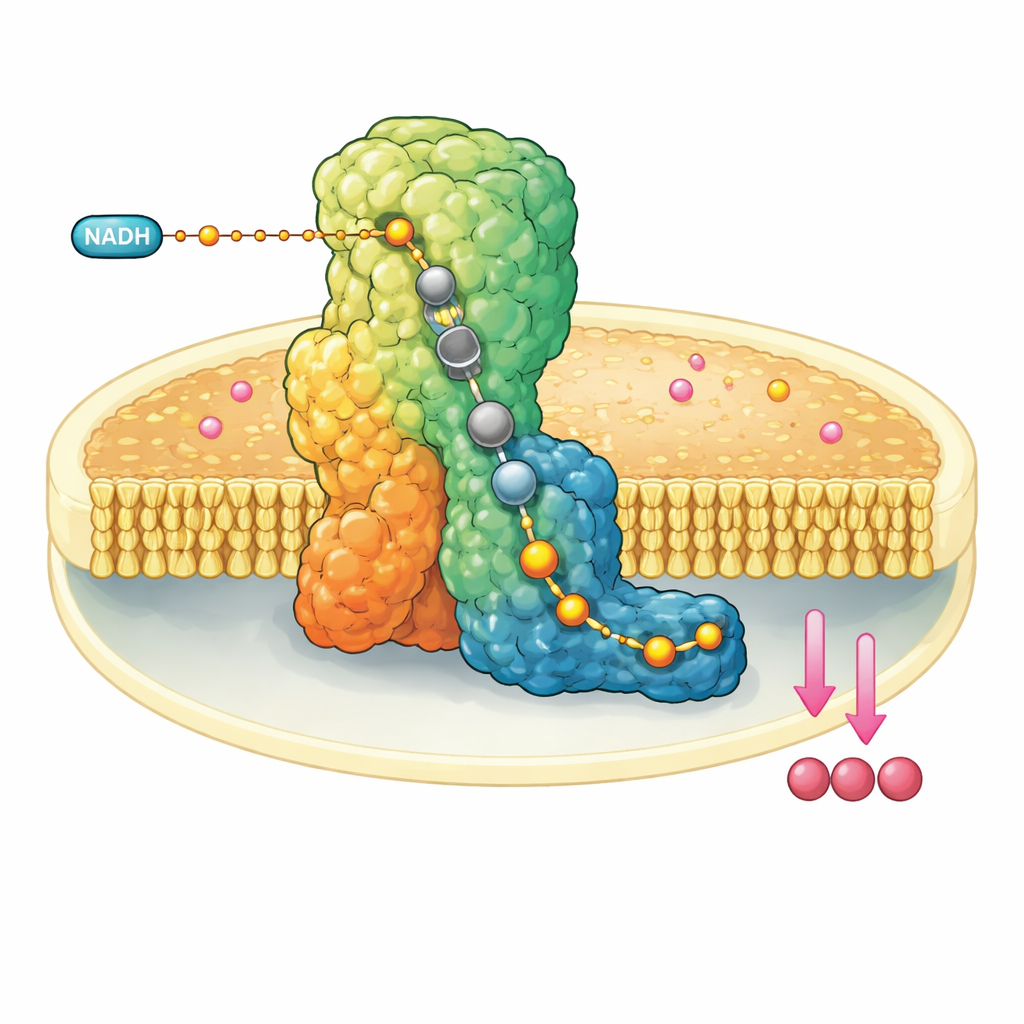
Une machine moléculaire dans la paroi mitochondriale
Le complexe I est un énorme assemblage protéique en forme de L inséré dans la membrane interne des mitochondries. Il prélève des électrons d’une molécule combustible, le NADH, et les transmet le long d’un fil intégré de clusters métalliques jusqu’à l’ubiquinone‑10, un transporteur mobile qui relaie ensuite les électrons le long de la chaîne respiratoire. Parallèlement, le complexe I utilise l’énergie libérée pour déplacer quatre protons à travers la membrane, contribuant à établir un gradient électrochimique qui alimente en fin de compte la synthèse d’ATP. En raison de son rôle central, un dysfonctionnement du complexe I est lié à diverses pathologies neuromusculaires et métaboliques ainsi qu’à des lésions tissulaires lors d’une interruption de l’apport sanguin.
Geler la machine en plein souffle
Les chercheurs ont reconstitué le complexe I bovin dans de petits disques de membrane artificielle contenant également de l’ubiquinone‑10. Ils ont ensuite mélangé rapidement un grand excès de NADH et plongé l’échantillon dans l’azote liquide en quelques dizaines de secondes, piégeant l’enzyme après qu’elle ait effectué au moins un cycle catalytique. Grâce à la cryo‑microscopie électronique de pointe à des résolutions descendant jusqu’à environ deux angströms — suffisantes pour distinguer des molécules d’eau individuelles — ils ont pu reconstruire plusieurs structures tridimensionnelles du complexe I dans différents états au repos. De façon cruciale, dans la forme dite fermée et active, ils ont observé l’ubiquinol‑10, le produit réduit, occupant entièrement la longueur du canal étroit où se déroule la réaction.
Suivre le trajet du vecteur d’énergie
Dans la forme active, l’équipe a observé deux conformations distinctes de la même molécule d’ubiquinol‑10 nichée à l’extrémité de son canal de liaison, près du cluster fer‑soufre terminal. Par rapport aux structures antérieures contenant l’ubiquinone oxydée, la tête de la molécule a changé de position et d’orientation, formant un fort lien hydrogène avec une chaîne latérale tyrosine particulière mais se trouvant trop éloignée d’une histidine voisine pour interagir directement. Des simulations informatiques complémentaires des charges environnantes et des molécules d’eau ont montré que cette configuration est la plus stable lorsque cette tyrosine et un résidu aspartate sont déprotonés — ce qui est cohérent avec le fait qu’ils viennent de céder les deux protons qui convertissent l’ubiquinone en ubiquinol. La seconde pose, moins bien définie, représente probablement une moyenne de plusieurs états produits étroitement liés alors que l’ubiquinol se prépare à quitter le site.
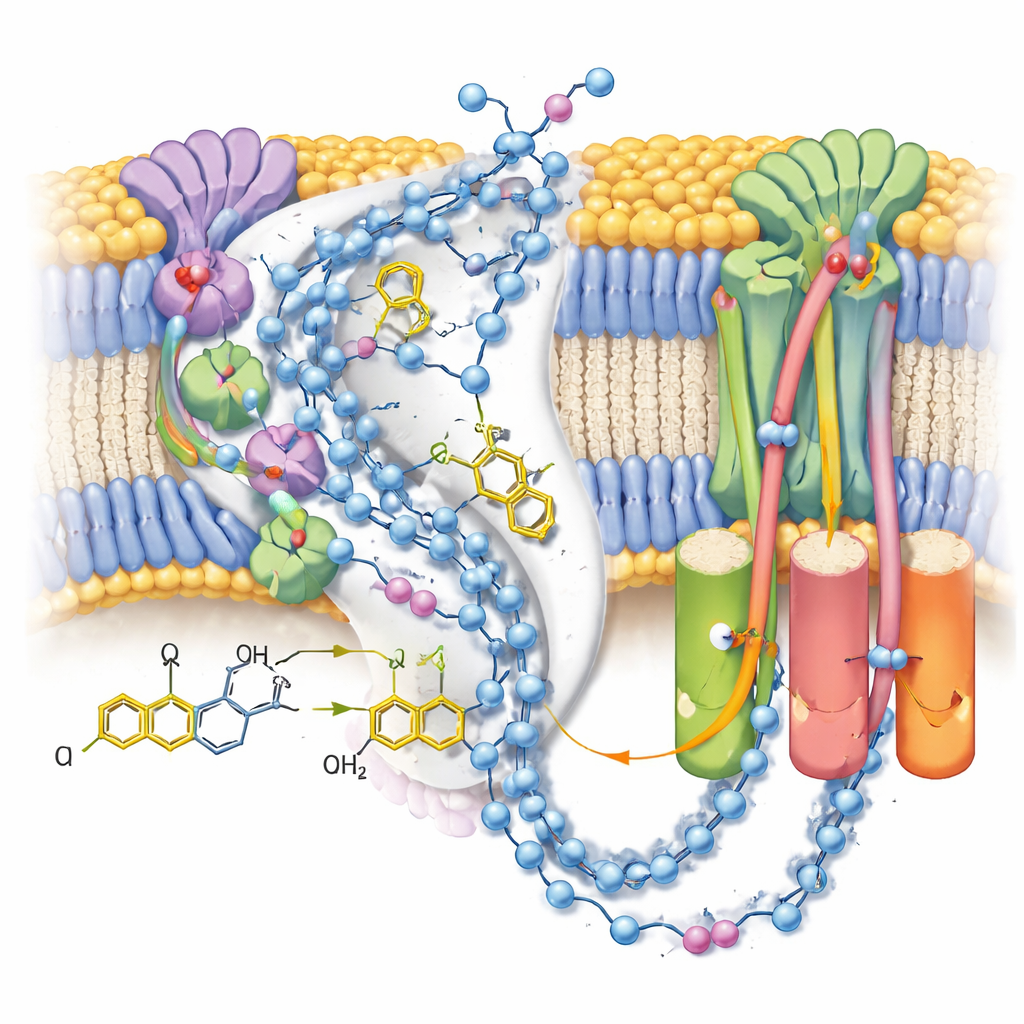
Cartographier les voies protoniques
Étant donné que les cartes ont résolu de nombreuses molécules d’eau ordonnées, les auteurs ont pu tracer des chaînes quasi continues de résidus polaires et d’eau à travers le bras membranaire du complexe I. Ces chaînes relient le site de réaction de l’ubiquinone à trois modules séparés « de type anti‑porteuse » qui déplacent des protons à travers la membrane. Dans l’état fermé post‑réaction avec l’ubiquinol lié, les connexions semblent largement intactes mais ponctuées de petits intervalles où l’eau est vraisemblablement trop mobile pour être observée, ce qui suggère des voies flexibles plutôt que des conduits rigides. En revanche, un état alternatif ouvert et désactivé — stabilisé ici par une molécule de détergent logée dans le canal — montre une hélice cintrée qui rompt une connexion clé, soutenant l’idée que cet état régulateur bloque temporairement le transfert de protons sur longue distance.
Ce que cela signifie pour la santé cellulaire
Pris ensemble, les clichés structuraux et les simulations étayent une séquence dans laquelle l’ubiquinone se lie dans une pose pré‑réaction, accepte deux électrons et deux protons pour devenir ubiquinol, puis persiste dans une pose post‑réaction pendant que les groupes protéiques voisins attendent d’être reprotonés du côté matrice de la membrane. Le travail clarifie comment de subtils changements de position et de charge de quelques atomes au site actif peuvent se propager le long de voies protoniques étendues pour entraîner le pompage, et comment des formes alternatives désactivées peuvent interrompre ce flux. En affinant notre vision du complexe I en action, ces résultats fournissent un cadre pour comprendre les défauts héréditaires, l’action des médicaments et les lésions mitochondriales qui surviennent lors d’événements tels que les infarctus et les AVC.
Citation: Chung, I., Pereira, C.S., Wright, J.J. et al. Post-catalysis structures of mitochondrial complex I with ubiquinol-10 bound in the active site. Nat Commun 17, 3506 (2026). https://doi.org/10.1038/s41467-026-70030-0
Mots-clés: complexe I mitochondrial, ubiquinone‑10, structure cryo‑EM, pompage de protons, phosphorylation oxydative