Clear Sky Science · pl
Struktury po katalizie kompleksu I mitochondrium z ubiquinolem-10 związanym w miejscu aktywnym
Jak nasze komórki zamieniają pokarm w paliwo
Mitochondria często nazywane są elektrowniami komórek, a jednym z ich kluczowych zespołów, znanym jako kompleks I, rozpoczyna proces przekształcania energii z pożywienia w cząsteczkę ATP. W tym badaniu przyjrzano się kompleksowi I z dokładnością atomową, aby zobaczyć, jak dokładnie wygląda tuż po wykonaniu swojej pracy, kiedy mała lipidopodobna cząsteczka zwana ubichinonem-10 została właśnie przekształcona w swoją naładowaną, bogatą w energię formę. Zrozumienie tej ulotnej chwili pomaga wyjaśnić, jak komórki tak efektywnie wychwytują energię i dlaczego zaburzenia tego mechanizmu mogą przyczyniać się do chorób mózgu, mięśni i serca.
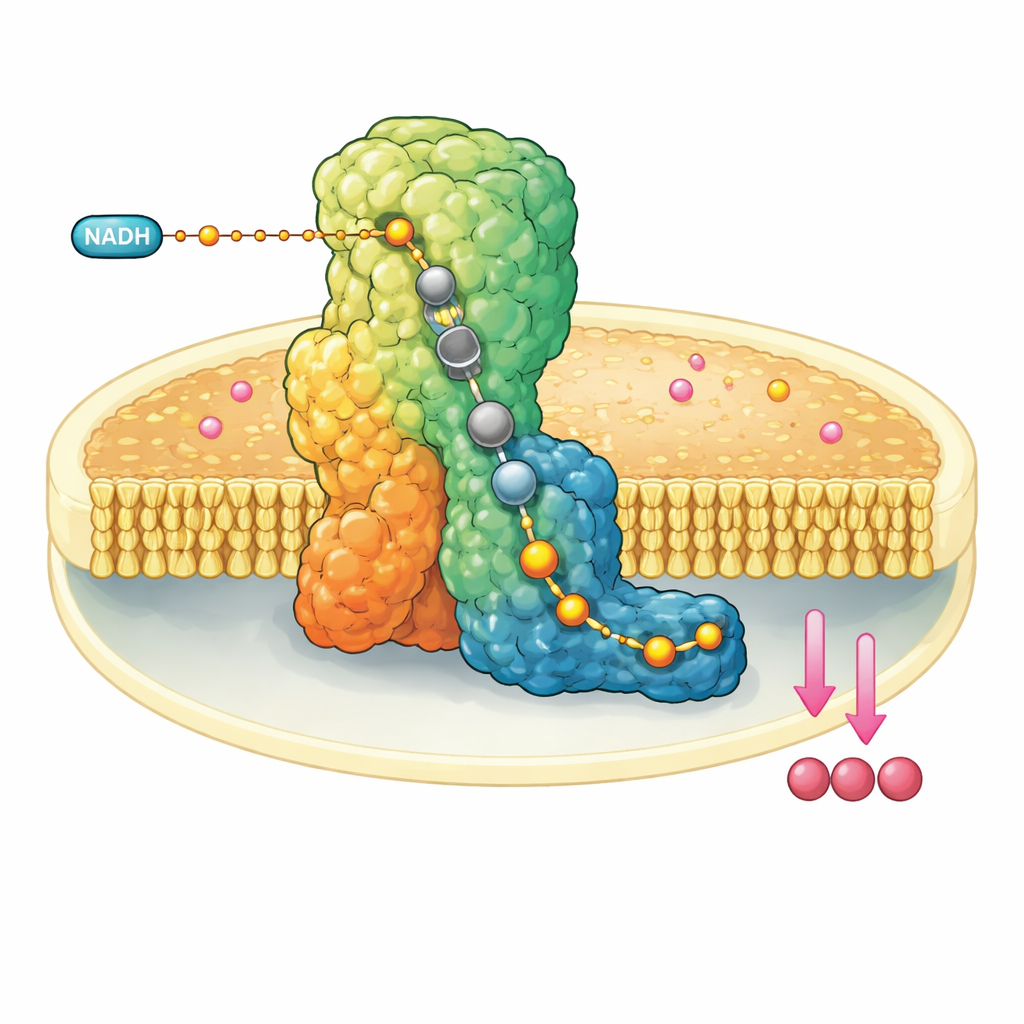
Molekularna maszyna w ścianie mitochondrium
Kompleks I to ogromny, w kształcie litery L zespół białkowy osadzony w wewnętrznej błonie mitochondriów. Pobiera elektrony z cząsteczki paliwa, NADH, i przekazuje je w dół wbudowanego „przewodu” z klastrów metaloorganicznych, aż trafiają do ubichinonu-10, ruchomego nośnika, który dalej transportuje elektrony w łańcuchu oddechowym. Równocześnie kompleks I wykorzystuje uwolnioną energię do przeniesienia czterech protonów przez błonę, pomagając zbudować gradient elektrochemiczny, który ostatecznie napędza syntezę ATP. Ze względu na centralną rolę, dysfunkcja kompleksu I wiąże się z szeregiem zaburzeń nerwowo-mięśniowych i metabolicznych oraz z uszkodzeniem tkanek podczas przerwanego dopływu krwi.
Zamrażanie maszyny w połowie „oddechu”
Naukowcy rekonstruowali byczy kompleks I w maleńkich sztucznych dyskach błonowych, które zawierały także ubichinon-10. Następnie szybko wymieszali dużą nadwyżkę NADH i błyskawicznie zamrozili próbkę w ciągu kilkudziesięciu sekund, uwięzając enzym po wykonaniu co najmniej jednego cyklu katalitycznego. Korzystając z najnowocześniejszej krioelektronowej mikroskopii elektronowej o rozdzielczościach sięgających około dwóch angstremów — wystarczająco, aby zobaczyć pojedyncze cząsteczki wody — mogli odtworzyć wiele trójwymiarowych struktur kompleksu I w różnych stanach spoczynkowych. Co istotne, w tak zwanej zamkniętej, aktywnej formie zaobserwowano ubiquinol-10, zredukowany produkt, zajmujący w pełni całą długość wąskiego kanału, w którym zachodzi reakcja.
Śledzenie drogi nośnika energii
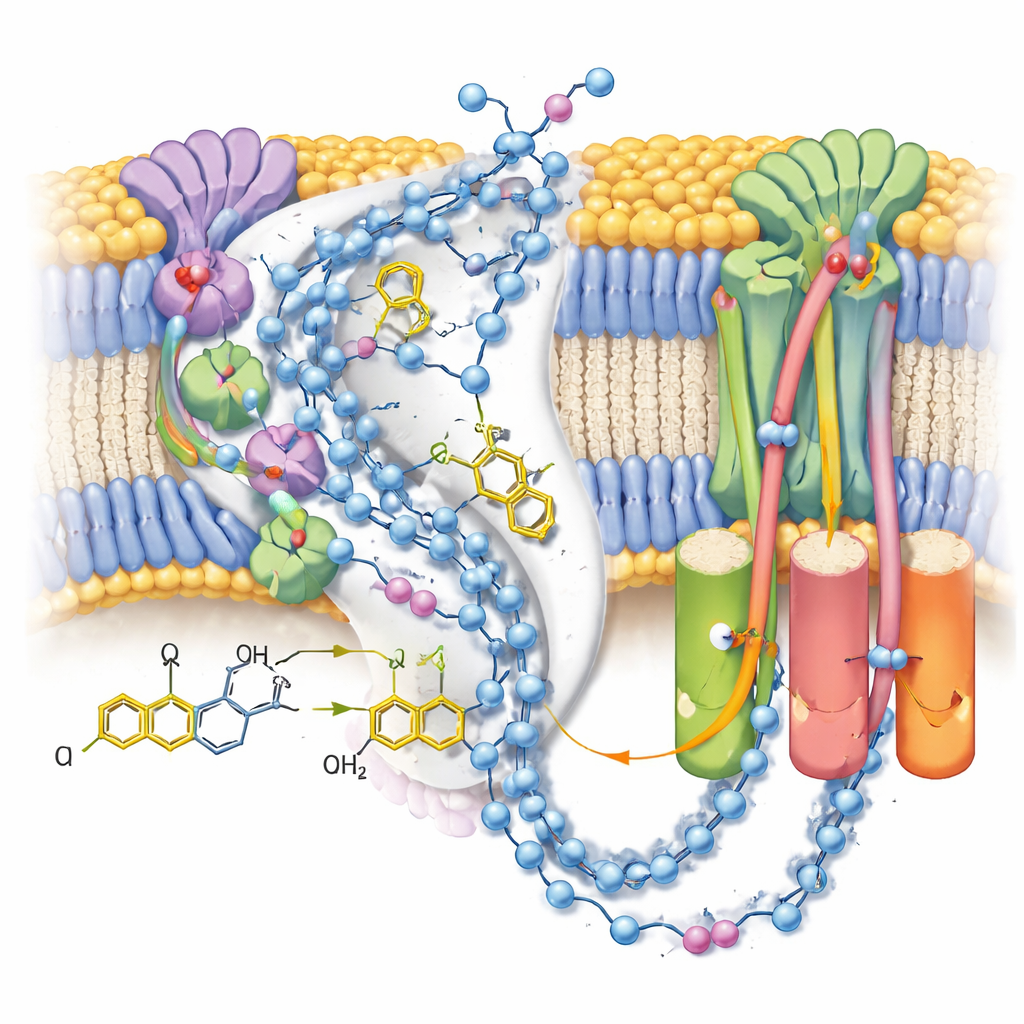
W formie aktywnej zespół zaobserwował dwie odrębne konformacje tej samej cząsteczki ubiquinolu-10 ułożone na końcu jej kanału wiążącego, blisko końcowego klastra żelazo-siarkowego. W porównaniu z wcześniejszymi strukturami zawierającymi utleniony ubichinon, główka cząsteczki przesunęła pozycję i orientację, tworząc silne wiązanie wodorowe z określonym łańcuchem bocznym tyrozyny, ale znajdując się zbyt daleko od pobliskiej histydyny, by bezpośrednio z nią oddziaływać. Uzupełniające symulacje komputerowe rozkładów ładunków i cząsteczek wody pokazały, że ta konfiguracja jest najbardziej stabilna, gdy zarówno ta tyrozyna, jak i reszta asparaginianu są zdeprotonowane — co jest zgodne z tym, że właśnie oddały dwa protony przekształcające ubichinon w ubiquinol. Druga, słabiej zdefiniowana pozycja prawdopodobnie reprezentuje średnią z kilku blisko spokrewnionych stanów produktu, gdy ubiquinol przygotowuje się do opuszczenia miejsca reakcji.
Przewodzenie szlaków protonowych
Dzięki mapom ukazującym wiele uporządkowanych cząsteczek wody autorzy mogli wyśledzić niemal ciągłe łańcuchy polarnych reszt i wody przez ramiona błonowe kompleksu I. Łańcuchy te łączą miejsce reakcji ubichinonu z trzema oddzielnymi modułami „podobnymi do antyporterów”, które przesuwają protony przez błonę. W stanie zamkniętym po reakcji z związanym ubiquinolem połączenia wydają się w dużej mierze nienaruszone, choć przerywane małymi szczelinami, gdzie woda prawdopodobnie jest zbyt ruchliwa, by ją zobaczyć, co sugeruje elastyczne ścieżki raczej niż sztywne rury. W przeciwieństwie do tego, alternatywny stan otwarty, nieaktywny — tutaj stabilizowany przez cząsteczkę detergentu zaklinowaną w kanale — wykazuje zgiętą helisę, która przerywa jedno kluczowe połączenie, co wspiera hipotezę, że ten regulacyjny stan tymczasowo blokuje długodystansowy transfer protonów.
Co to oznacza dla zdrowia komórek
W skrócie, migawki strukturalne i symulacje wspierają sekwencję, w której ubichinon wiąże się w pozycji przedreakcyjnej, przyjmuje dwa elektrony i dwa protony stając się ubiquinolem, a następnie pozostaje w pozycji posreakcyjnej, podczas gdy pobliskie grupy białkowe czekają na ponowne protonowanie z strony macierzy mitochondrialnej. Praca wyjaśnia, jak subtelne zmiany w położeniu i ładunku kilku atomów w miejscu aktywnym mogą rozprzestrzeniać się przez długie ścieżki protonowe, napędzając pompowanie, oraz jak alternatywne, nieaktywne formy mogą przerywać ten przepływ. Poprzez wyostrzenie naszego obrazu kompleksu I w działaniu, wyniki te dostarczają ram do zrozumienia wrodzonych wad, działania leków oraz uszkodzeń mitochondriów, które występują podczas zdarzeń takich jak zawały serca i udary.
Cytowanie: Chung, I., Pereira, C.S., Wright, J.J. et al. Post-catalysis structures of mitochondrial complex I with ubiquinol-10 bound in the active site. Nat Commun 17, 3506 (2026). https://doi.org/10.1038/s41467-026-70030-0
Słowa kluczowe: kompleks I mitochondriów, ubichinon-10, struktura cryo-EM, pompa protonowa, fosforylacja oksydacyjna