Clear Sky Science · es
Estructuras post-catalíticas del complejo I mitocondrial con ubiquinol-10 unido en el sitio activo
Cómo nuestras células convierten los alimentos en combustible
Las mitocondrias suelen llamarse las centrales energéticas de nuestras células, y una de sus máquinas clave, conocida como complejo I, inicia el proceso de transformar la energía de los alimentos en la molécula ATP. Este estudio se aproxima al complejo I con detalle atómico para ver exactamente cómo es justo después de haber hecho su trabajo, cuando una pequeña molécula lipídica llamada ubiquinona-10 acaba de convertirse en su forma cargada y rica en energía. Entender este momento fugaz ayuda a explicar cómo nuestras células capturan la energía con tanta eficiencia y por qué las alteraciones en esta maquinaria pueden contribuir a enfermedades del cerebro, los músculos y el corazón.
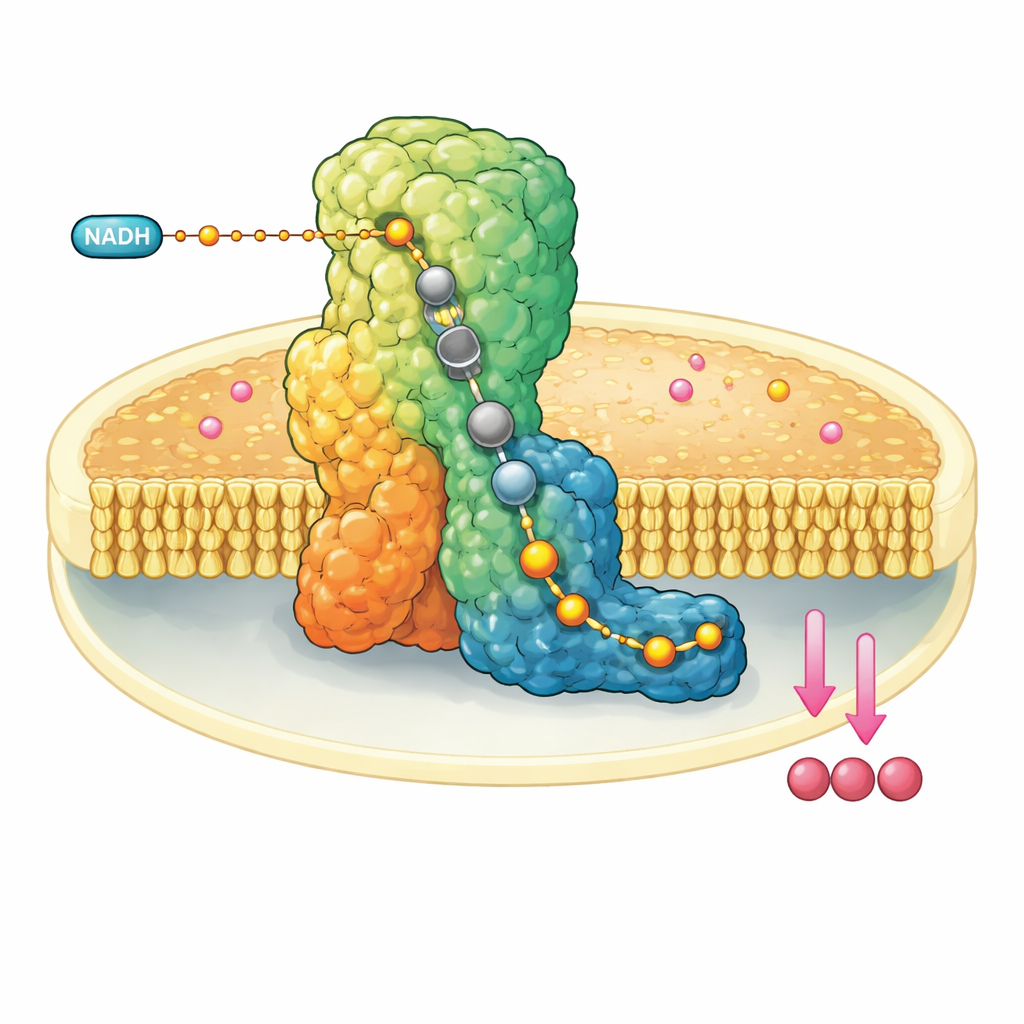
Una máquina molecular en la pared mitocondrial
El complejo I es un enorme ensamblaje proteico con forma de L incrustado en la membrana interna de las mitocondrias. Extrae electrones de una molécula combustible, el NADH, y los pasa por un cable interno de clústeres metálicos hasta que alcanzan la ubiquinona-10, un transportador móvil que lleva electrones más adelante en la cadena respiratoria. Al mismo tiempo, el complejo I utiliza la energía liberada para mover cuatro protones a través de la membrana, contribuyendo a la creación de un gradiente electroquímico que finalmente impulsa la síntesis de ATP. Debido a su papel central, el mal funcionamiento del complejo I se asocia con una serie de trastornos neuromusculares y metabólicos y con daño tisular durante interrupciones del suministro sanguíneo.
Congelando la máquina a mitad de respiración
Los investigadores reconstituyeron el complejo I bovino en diminutos discos de membrana artificial que también contenían ubiquinona-10. Luego mezclaron rápidamente un exceso grande de NADH y sumergieron la muestra en congelación rápida en cuestión de decenas de segundos, atrapando la enzima después de que hubiera realizado al menos un ciclo catalítico. Usando microscopía crioelectrónica de última generación a resoluciones de hasta alrededor de dos angstroms —suficiente para ver moléculas de agua individuales— pudieron reconstruir múltiples estructuras tridimensionales del complejo I en diferentes formas de reposo. De forma crucial, en la llamada forma cerrada y activa, observaron ubiquinol-10, el producto reducido, ocupando completamente toda la longitud del estrecho canal donde tiene lugar la reacción.
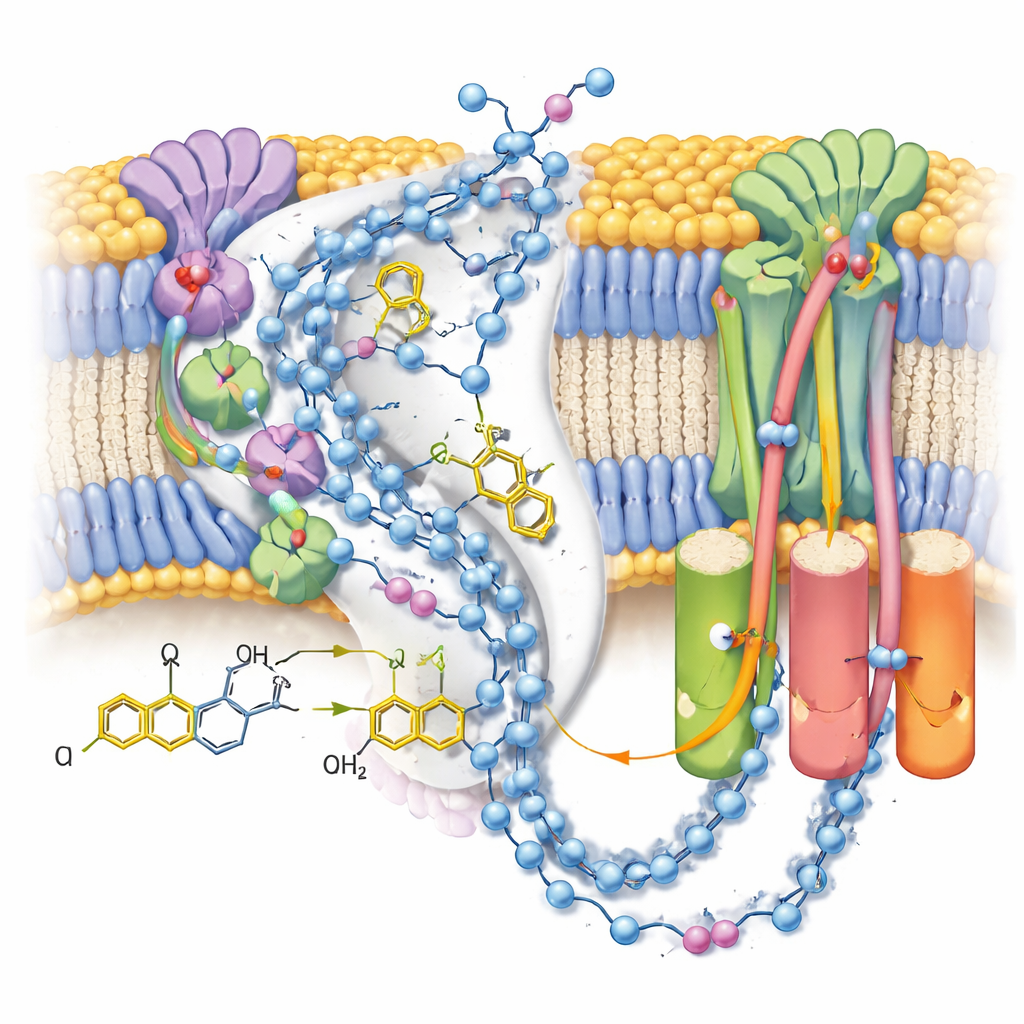
Siguiendo la senda del transportador de energía
En la forma activa, el equipo observó dos conformaciones distintas de la misma molécula de ubiquinol-10 alojadas al final de su canal de unión cerca del clúster hierro-azufre terminal. En comparación con estructuras previas con ubiquinona oxidada, la cabeza de la molécula había cambiado de posición y orientación, formando un fuerte enlace de hidrógeno con la cadena lateral de una tirosina concreta pero quedando demasiado lejos de una histidina cercana para interactuar directamente. Simulaciones informáticas complementarias de las cargas circundantes y las moléculas de agua mostraron que esta configuración es más estable cuando tanto esa tirosina como un residuo de aspartato están desprotonados —consistente con que acaban de donar los dos protones que convierten ubiquinona en ubiquinol. La segunda pose, más débilmente definida, probablemente representa un promedio de varios estados de producto estrechamente relacionados mientras el ubiquinol se prepara para abandonar el sitio.
Explorando las vías de protones
Dado que los mapas resolvieron muchas moléculas de agua ordenadas, los autores pudieron trazar cadenas casi continuas de residuos polares y agua a través del brazo de la membrana del complejo I. Estas cadenas conectan el sitio de la reacción de la ubiquinona con tres módulos separados «tipo antiportador» que mueven protones a través de la membrana. En el estado cerrado post-reacción con ubiquinol unido, las conexiones parecen en gran medida intactas pero salpicadas de pequeños huecos donde el agua probablemente es demasiado móvil para observarse, lo que sugiere vías flexibles más que tuberías rígidas. En contraste, un estado alternativo abierto y desactivado —estabilizado aquí por una molécula detergente alojada en el canal— muestra una hélice doblada que rompe una conexión clave, apoyando la idea de que este estado regulador bloquea temporalmente la transferencia de protones a larga distancia.
Qué significa esto para la salud celular
En conjunto, las instantáneas estructurales y las simulaciones respaldan una secuencia en la que la ubiquinona se une en una pose previa a la reacción, acepta dos electrones y dos protones para convertirse en ubiquinol, y luego permanece en una pose post-reacción mientras los grupos proteicos cercanos esperan ser reprotonados desde el lado de la matriz de la membrana. El trabajo aclara cómo cambios sutiles en la posición y la carga de unos pocos átomos en el sitio activo pueden propagarse a lo largo de largas vías de protones para impulsar el bombeo, y cómo formas alternativas desactivadas pueden interrumpir este flujo. Al afinar nuestra visión del complejo I en funcionamiento, estos resultados proporcionan un marco para entender defectos heredados, la acción de fármacos y el daño mitocondrial que ocurre durante eventos como infartos y accidentes cerebrovasculares.
Cita: Chung, I., Pereira, C.S., Wright, J.J. et al. Post-catalysis structures of mitochondrial complex I with ubiquinol-10 bound in the active site. Nat Commun 17, 3506 (2026). https://doi.org/10.1038/s41467-026-70030-0
Palabras clave: complejo I mitocondrial, ubiquinona-10, estructura por crio-EM, bombeo de protones, fosforilación oxidativa