Clear Sky Science · de
Post-katalytische Strukturen des mitochondrialen Komplex I mit an der aktiven Stelle gebundenem Ubiquinol-10
Wie unsere Zellen Nahrung in Treibstoff verwandeln
Mitochondrien werden oft als Kraftwerke unserer Zellen bezeichnet, und eine ihrer Schlüsselmaschinen, bekannt als Komplex I, leitet den Prozess ein, mit dem die in unserer Nahrung gespeicherte Energie in das Molekül ATP überführt wird. Diese Studie zoomt auf Komplex I auf atomare Details, um genau zu sehen, wie er unmittelbar nach seiner Arbeit aussieht, wenn ein kleines fettähnliches Molekül namens Ubiquinon-10 gerade in seine geladene, energiereiche Form umgewandelt wurde. Das Verständnis dieses flüchtigen Moments hilft zu erklären, wie unsere Zellen so effizient Energie einfangen und warum Störungen in dieser Maschinerie zu Erkrankungen des Gehirns, der Muskeln und des Herzens beitragen können.
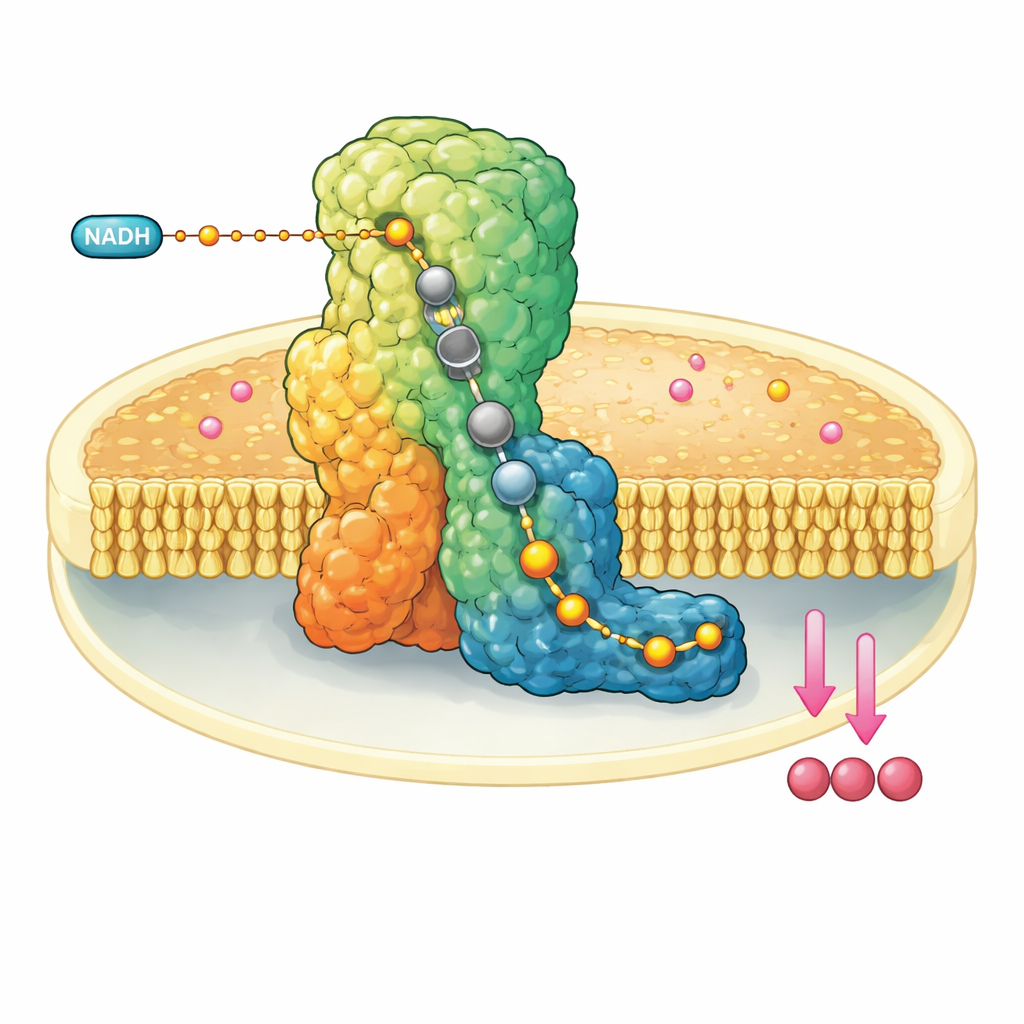
Eine molekulare Maschine in der Mitochondrienmembran
Komplex I ist ein riesiger, L-förmiger Proteinkomplex, der in die innere Membran der Mitochondrien eingebettet ist. Er entnimmt Elektronen von einem Brennstoffmolekül, NADH, und leitet sie über eine eingebaute Leitung aus Metallclustern weiter, bis sie Ubiquinon-10 erreichen, einem mobilen Carrier, der die Elektronen entlang der Atmungskette weitertransportiert. Gleichzeitig nutzt Komplex I die freigesetzte Energie, um vier Protonen über die Membran zu befördern und so ein elektrochemisches Gefälle aufzubauen, das letztlich die ATP-Synthese antreibt. Wegen seiner zentralen Rolle ist eine Fehlfunktion von Komplex I mit einer Reihe neuromuskulärer und metabolischer Störungen sowie mit Gewebeschäden bei unterbrochener Blutversorgung verbunden.
Die Maschine mitten im Atemzug einfrieren
Die Forscher rekonstruierten bovines Komplex I in winzige künstliche Membranscheiben, die ebenfalls Ubiquinon-10 enthielten. Sie mischten dann schnell einen großen Überschuss an NADH dazu und tauchten die Probe innerhalb weniger Dutzend Sekunden ein, um das Enzym nach mindestens einem katalytischen Zyklus einzufangen. Mit hochmoderner Kryo-Elektronenmikroskopie bei Auflösungen bis etwa zwei Angström — genug, um einzelne Wassermoleküle zu sehen — konnten sie mehrere dreidimensionale Strukturen von Komplex I in verschiedenen Ruheformen rekonstruieren. Entscheidend ist, dass sie in der sogenannten geschlossenen, aktiven Form Ubiquinol-10, das reduzierte Produkt, beobachteten, das den gesamten schmalen Kanal, in dem die Reaktion stattfindet, vollständig besetzt.
Den Weg des Energieträgers verfolgen
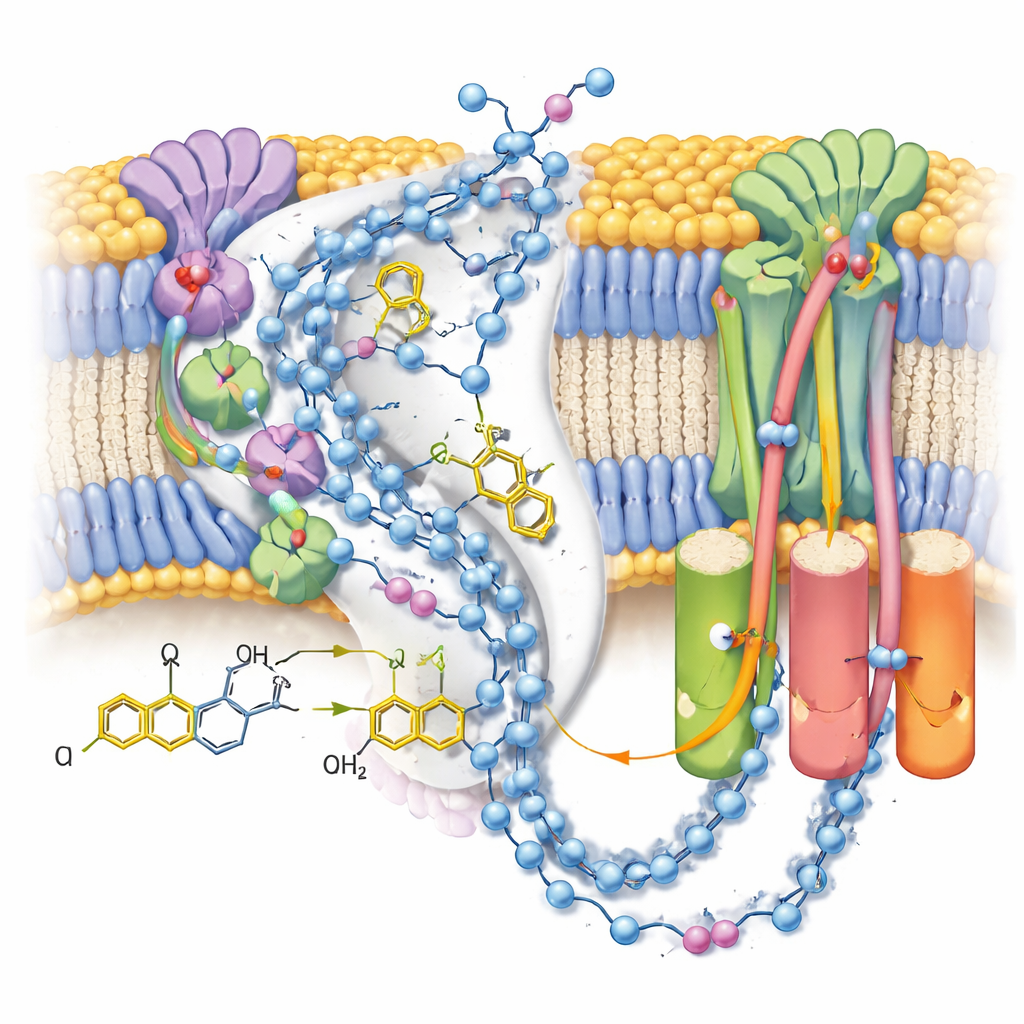
In der aktiven Form sah das Team zwei unterschiedliche Konformationen desselben Ubiquinol-10-Moleküls, eingebettet am Ende seines Bindungskanals in der Nähe des terminalen Eisen–Schwefel-Clusters. Verglichen mit früheren Strukturen mit oxidiertem Ubiquinon hatte sich der Kopf des Moleküls in Position und Orientierung verschoben, bildete eine starke Wasserstoffbrücke zu einer bestimmten Tyrosin-Seitenkette, lag aber zu weit von einem nahegelegenen Histidin entfernt, um direkt zu interagieren. Komplementäre Computersimulationen der umgebenden Ladungen und Wassermoleküle zeigten, dass diese Konfiguration am stabilsten ist, wenn sowohl jenes Tyrosin als auch eine Aspartat-Rest deprotoniert sind — konsistent damit, dass sie gerade die beiden Protonen gespendet haben, die Ubiquinon in Ubiquinol verwandeln. Die zweite, schwächer definierte Pose stellt wahrscheinlich einen Durchschnitt mehrerer eng verwandter Produktzustände dar, während sich Ubiquinol auf das Verlassen der Bindungsstelle vorbereitet.
Die Protonenpfade erkunden
Da die Karten viele geordnete Wassermoleküle auflösten, konnten die Autoren nahezu kontinuierliche Ketten polarer Reste und Wassermoleküle durch den Membranarm von Komplex I zurückverfolgen. Diese Ketten verbinden die Ubiquinon-Reaktionsstelle mit drei getrennten „Antiporter‑ähnlichen“ Modulen, die Protonen über die Membran transportieren. Im postreaktiven, geschlossenen Zustand mit gebundenem Ubiquinol erscheinen die Verbindungen weitgehend intakt, sind jedoch von kleinen Lücken durchsetzt, in denen Wasser wahrscheinlich zu mobil ist, um sichtbar zu sein — ein Hinweis auf flexible Pfade statt starrer Rohre. Im Gegensatz dazu zeigt ein alternativer offener, deaktivierter Zustand — hier durch ein im Kanal eingeklemmtes Detergens stabilisiert — eine geknickte Helix, die eine wichtige Verbindung unterbricht und die Idee stützt, dass dieser regulatorische Zustand vorübergehend den langreichweitigen Protonentransfer blockiert.
Was das für die Zellgesundheit bedeutet
Insgesamt stützen die strukturellen Momentaufnahmen und Simulationen eine Abfolge, in der Ubiquinon in einer Vorreaktionslage bindet, zwei Elektronen und zwei Protonen aufnimmt, um zu Ubiquinol reduziert zu werden, und dann in einer Nachreaktionslage verweilt, während die nahegelegenen Proteingruppen darauf warten, von der Matrixseite der Membran wieder protoniert zu werden. Die Arbeit klärt, wie subtile Änderungen in Position und Ladung weniger Atome an der aktiven Stelle über lange Protonenpfade hinweg die Pumpleistung antreiben können und wie alternative, deaktivierte Formen diesen Fluss unterbrechen. Indem sie unseren Blick auf Komplex I in Aktion schärfen, liefern diese Ergebnisse einen Rahmen zum Verständnis vererbter Defekte, der Wirkung von Arzneimitteln und des mitochondrialen Schadens, der bei Ereignissen wie Herzinfarkten und Schlaganfällen auftritt.
Zitation: Chung, I., Pereira, C.S., Wright, J.J. et al. Post-catalysis structures of mitochondrial complex I with ubiquinol-10 bound in the active site. Nat Commun 17, 3506 (2026). https://doi.org/10.1038/s41467-026-70030-0
Schlüsselwörter: mitochondrialer Komplex I, Ubiquinon-10, cryo-EM-Struktur, Protonenpumpen, oxidative Phosphorylierung