Clear Sky Science · nl
Structuren na katalyse van mitochondriaal complex I met ubiquinol-10 gebonden in de actieve plaats
Hoe onze cellen voedsel in brandstof omzetten
Mitochondriën worden vaak de energiecentrales van onze cellen genoemd, en een van hun belangrijkste machines, bekend als complex I, start het proces waarbij de energie uit ons voedsel wordt omgezet in het molecuul ATP. Deze studie zoemt in op complex I op atomaire schaal om precies te zien hoe het eruitziet net nadat het zijn werk heeft gedaan, wanneer een klein vetachtig molecuul genaamd ubiquinon-10 zojuist is omgezet in zijn geladen, energierijke vorm. Inzicht in dit vluchtige moment helpt verklaren hoe onze cellen energie zo efficiënt vastleggen en waarom verstoringen in dit apparaat kunnen bijdragen aan ziekten van de hersenen, spieren en het hart.
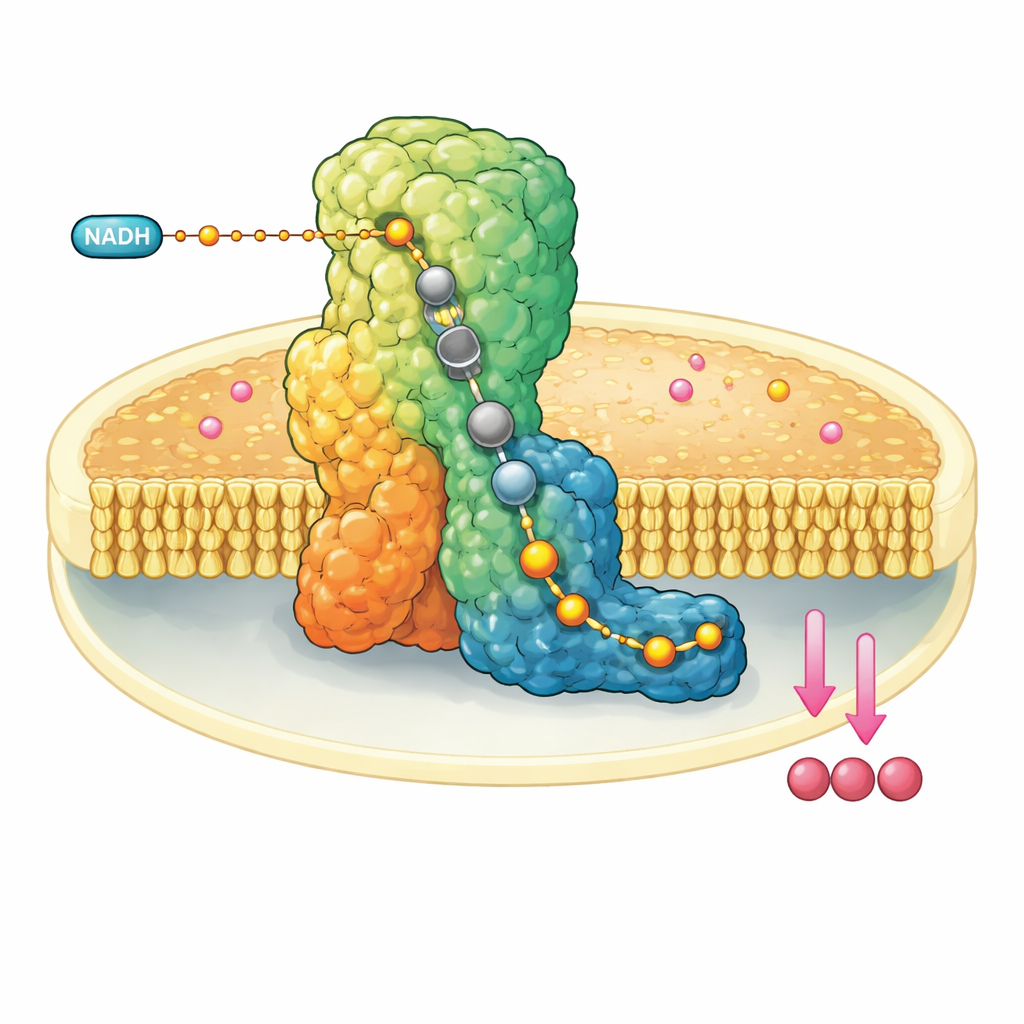
Een moleculaire machine in de mitochondriale wand
Complex I is een enorm L-vormig eiwitcomplex ingebed in het binnenste membraan van mitochondriën. Het onttrekt elektronen aan een brandstofmolecuul, NADH, en geeft ze door langs een ingebouwde draad van metaalclusters totdat ze ubiquinon-10 bereiken, een mobiele drager die elektronen verder langs de ademhalingsketen transporteert. Tegelijk gebruikt complex I de vrijgekomen energie om vier protonen over het membraan te verplaatsen, wat bijdraagt aan de opbouw van een elektrochemische gradiënt die uiteindelijk de ATP-synthese aandrijft. Vanwege zijn centrale rol wordt een disfunctie van complex I in verband gebracht met een reeks neuromusculaire en stofwisselingsstoornissen en met weefselschade tijdens onderbroken bloedtoevoer.
De machine bevriezen halverwege een ademhaling
De onderzoekers reconstrueerden runds complex I in kleine kunstmatige membraanschijven die ook ubiquinon-10 bevatten. Ze voegden vervolgens snel een grote overmaat aan NADH toe en vroren het monster binnen enkele tientallen seconden in, waardoor het enzym werd vastgelegd nadat het ten minste één katalytische cyclus had doorlopen. Met state-of-the-art cryo-elektronenmicroscopie bij resoluties tot ongeveer twee angstroom—genoeg om afzonderlijke watermoleculen te zien—konden ze meerdere driedimensionale structuren van complex I in verschillende rustvormen reconstrueren. Cruciaal was dat ze in de zogenoemde gesloten, actieve vorm ubiquinol-10, het gereduceerde product, observeerden dat de gehele lengte van het smalle kanaal waarin de reactie plaatsvond volledig vulde.
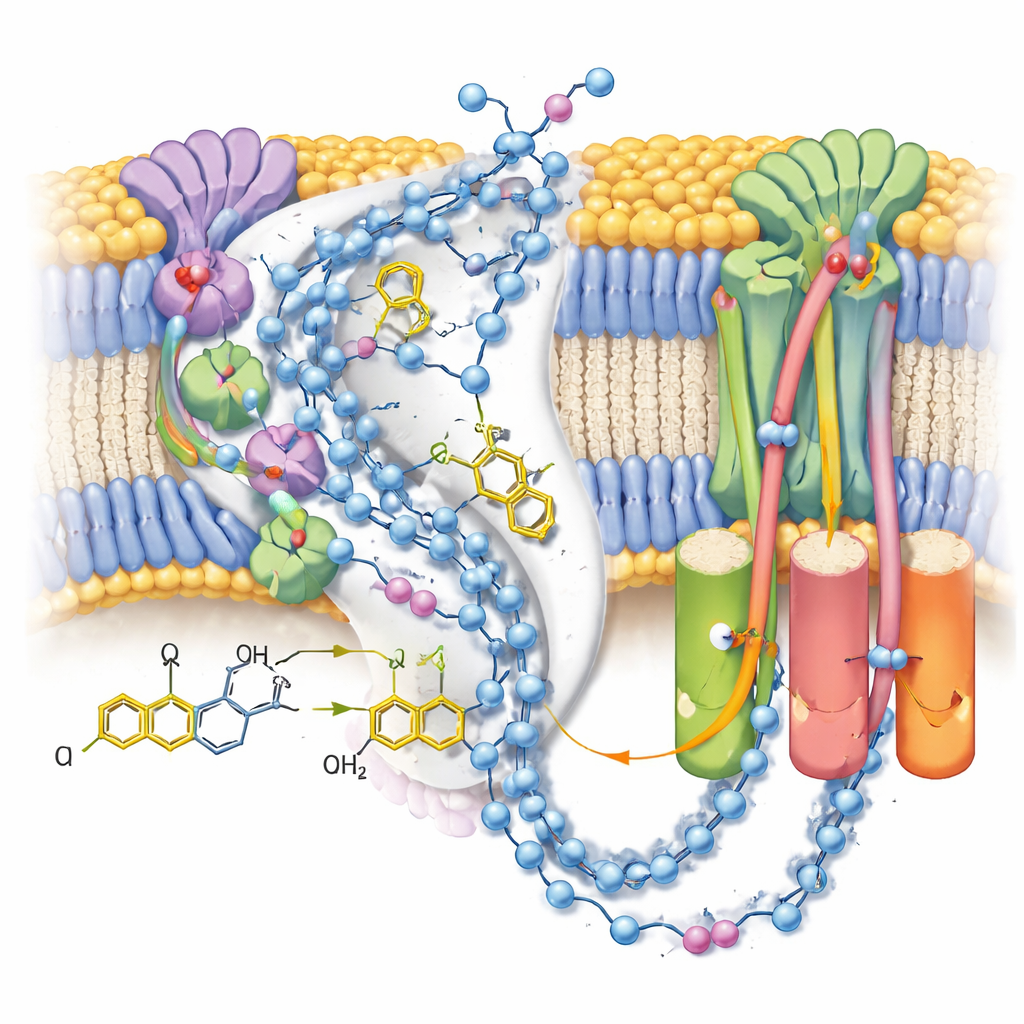
Het pad van de energiedrager volgen
In de actieve vorm zag het team twee verschillende conformaties van hetzelfde ubiquinol-10-molecuul, genesteld aan het einde van zijn bindingskanaal nabij de terminale ijzer‑zwavelcluster. Vergeleken met eerdere structuren met geoxideerd ubiquinon, had het kopgedeelte van het molecuul van positie en oriëntatie verschoven, en vormde het een sterke waterstofbinding met een specifieke tyrosine-zijgroep maar lag het te ver van een nabijgelegen histidine om direct te kunnen interageren. Complementaire computersimulaties van de omliggende ladingen en watermoleculen toonden aan dat deze configuratie het meest stabiel is wanneer zowel die tyrosine als een aspartaatresidu gedeprotoneerd zijn—wat overeenkomt met het feit dat ze net de twee protonen hebben gedoneerd die ubiquinon in ubiquinol omzetten. De tweede, zwakker gedefinieerde positie vertegenwoordigt waarschijnlijk een gemiddelde over verschillende nauw verwante producttoestanden terwijl ubiquinol zich voorbereidt om de plaats te verlaten.
De protonpaden in kaart brengen
Aangezien de kaarten veel geordende watermoleculen resolveerden, konden de auteurs bijna‑continue ketens van polaire residuen en water door de membraanarm van complex I volgen. Deze ketens verbinden de ubiquinon-reactieplaats met drie gescheiden “antiporter‑achtige” modules die protonen over het membraan verplaatsen. In de gesloten post‑reactietoestand met gebonden ubiquinol lijken de verbindingen grotendeels intact maar gepunctueerd door kleine gaps waar water waarschijnlijk te mobiel is om zichtbaar te zijn, wat wijst op flexibele paden in plaats van starre buizen. Ter vergelijking toont een alternatieve open, inactieve staat—hier gestabiliseerd door een detergentmolecuul dat in het kanaal was gekomen—een geknikte helix die een belangrijke verbinding verbreekt, wat het idee ondersteunt dat deze regulerende toestand tijdelijk langafstand‑protonoverdracht blokkeert.
Wat dit betekent voor de gezondheid van cellen
Samengevat ondersteunen de structurele momentopnamen en simulaties een volgorde waarin ubiquinon bindt in een pre‑reactiepose, twee elektronen en twee protonen accepteert om ubiquinol te worden, en vervolgens blijft hangen in een post‑reactiepose terwijl de nabijgelegen proteïnegroepen wachten om vanaf de matrixzijde van het membraan opnieuw geprotoneerd te worden. Het werk verduidelijkt hoe subtiele veranderingen in de positie en lading van een paar atomen in de actieve plaats zich kunnen voortplanten door lange protonpaden om pompwerking aan te sturen, en hoe alternatieve, inactieve vormen deze stroom kunnen onderbreken. Door ons beeld van complex I in werking te verscherpen, bieden deze resultaten een kader om erfelijke defecten, medicijnwerking en de mitochondriale schade die optreedt tijdens gebeurtenissen zoals hartaanvallen en beroertes te begrijpen.
Bronvermelding: Chung, I., Pereira, C.S., Wright, J.J. et al. Post-catalysis structures of mitochondrial complex I with ubiquinol-10 bound in the active site. Nat Commun 17, 3506 (2026). https://doi.org/10.1038/s41467-026-70030-0
Trefwoorden: mitochondriaal complex I, ubiquinon-10, cryo-EM-structuur, protonpomping, oxidatieve fosforylering