Clear Sky Science · it
Strutture post-catalisi del complesso I mitocondriale con ubichinolo-10 legato al sito attivo
Come le nostre cellule trasformano il cibo in carburante
I mitocondri sono spesso definiti centrali elettriche delle nostre cellule, e una delle loro macchine chiave, nota come complesso I, avvia il processo che converte l’energia del cibo nella molecola ATP. Questo studio mette a fuoco il complesso I a livello atomico per mostrare esattamente com’è appena dopo aver eseguito la sua funzione, quando una piccola molecola lipofila chiamata ubichinone-10 è stata appena convertita nella sua forma caricata e ricca di energia. Capire questo istante fugace aiuta a spiegare come le nostre cellule catturino l’energia con tanta efficienza e perché le alterazioni di questo apparato possano contribuire a malattie del cervello, dei muscoli e del cuore.
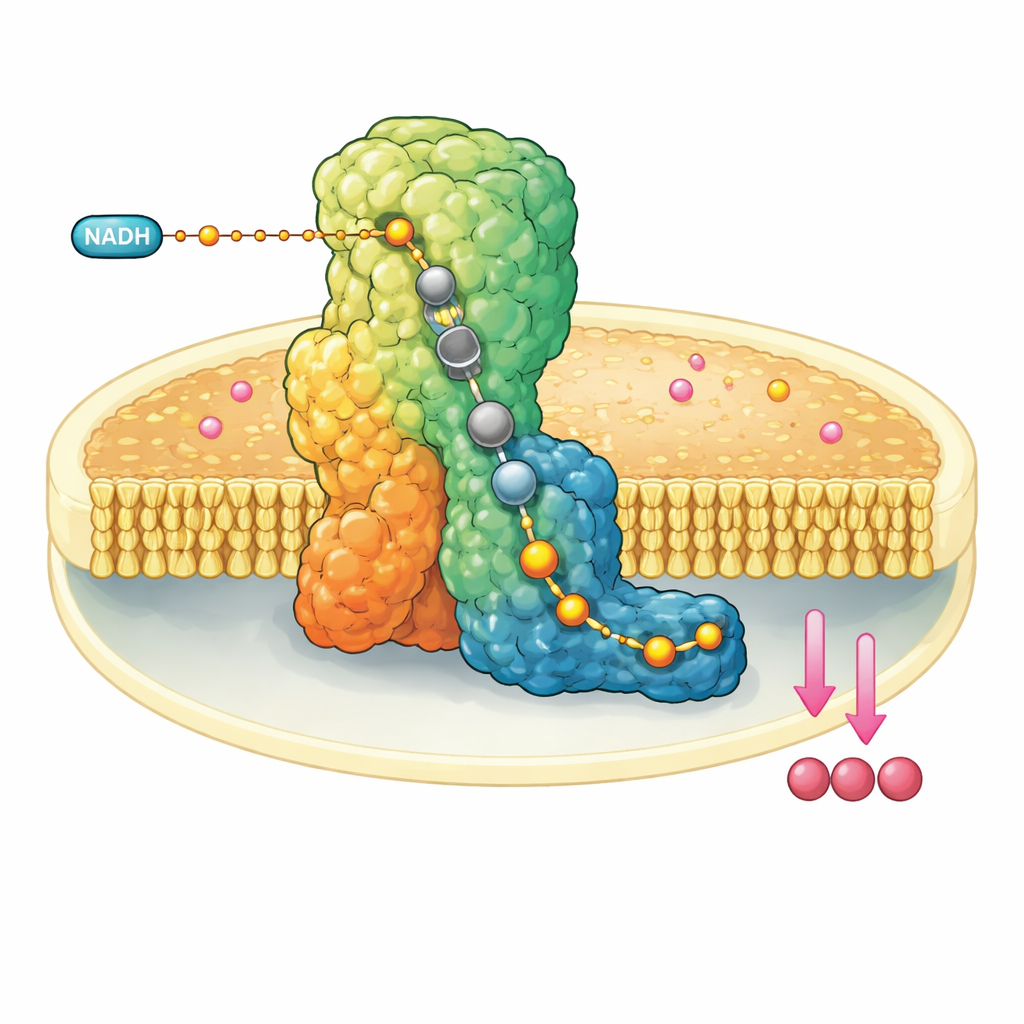
Una macchina molecolare nella parete mitocondriale
Il complesso I è un enorme assemblaggio proteico a forma di L incastrato nella membrana interna dei mitocondri. Preleva elettroni da una molecola di combustibile, il NADH, e li trasferisce lungo un filo interno formato da cluster metallici fino a raggiungere l’ubichinone-10, un trasportatore mobile che porta avanti gli elettroni nella catena respiratoria. Contemporaneamente, il complesso I usa l’energia rilasciata per spostare quattro protoni attraverso la membrana, contribuendo a costruire un gradiente elettrochimico che infine alimenta la sintesi di ATP. Per il suo ruolo centrale, il malfunzionamento del complesso I è collegato a una serie di disturbi neuromuscolari e metabolici e al danno tissutale durante l’interruzione del flusso sanguigno.
Congelare la macchina a metà respiro
I ricercatori hanno ricostituito il complesso I bovino in piccoli dischi di membrana artificiale che contenevano anche ubichinone-10. Hanno quindi miscelato rapidamente un grosso eccesso di NADH e congelato il campione per immersione nel giro di poche decine di secondi, intrappolando l’enzima dopo che aveva compiuto almeno un ciclo catalitico. Utilizzando microscopia crioelettronica all’avanguardia a risoluzioni fino a circa due angstrom—sufficienti per vedere singole molecole d’acqua—hanno potuto ricostruire più strutture tridimensionali del complesso I in diverse forme di riposo. Crucialmente, nella cosiddetta forma chiusa e attiva, hanno osservato l’ubichinolo-10, il prodotto ridotto, occupare completamente l’intera lunghezza del canale stretto dove avviene la reazione.
Seguire il percorso del trasportatore di energia
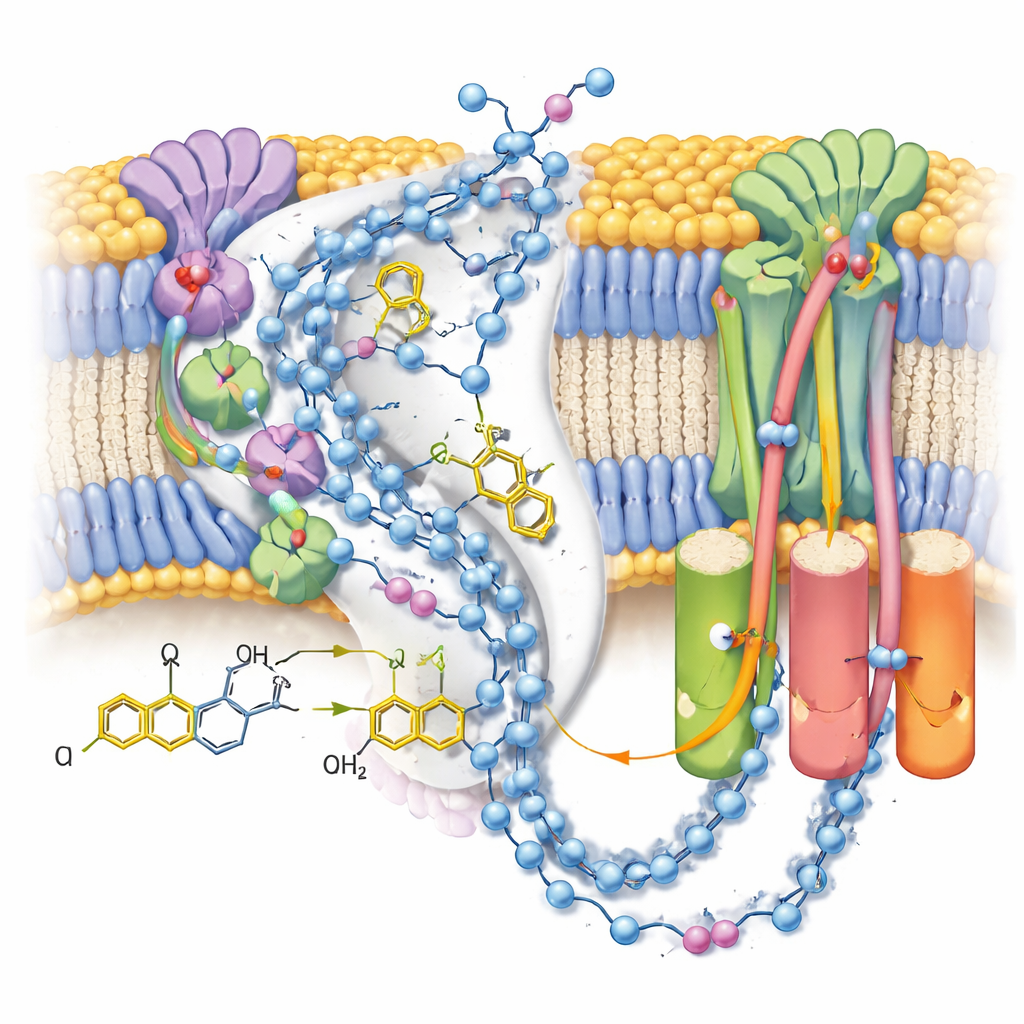
Nella forma attiva, il gruppo ha rilevato due conformazioni distinte della stessa molecola di ubichinolo-10 sistemata alla fine del suo canale di legame vicino al cluster ferro-zolfo terminale. Rispetto a strutture precedenti con l’ubichinone ossidato, la testa della molecola si era spostata in posizione e orientamento, formando un forte legame a idrogeno con una particolare catena laterale di tirosina ma trovandosi troppo distante da un istidina vicina per interagire direttamente. Simulazioni al computer complementari delle cariche circostanti e delle molecole d’acqua hanno mostrato che questa configurazione è più stabile quando sia quella tirosina sia un residuo aspartato sono deprotonati—coerente con il fatto che abbiano appena ceduto i due protoni che convertono l’ubichinone in ubichinolo. La seconda posa, meno definita, rappresenta probabilmente una media su diversi stati di prodotto strettamente correlati mentre l’ubichinolo si prepara a lasciare il sito.
Indagare le vie protoniche
Poiché le mappe hanno risolto molte molecole d’acqua ordinate, gli autori sono riusciti a tracciare catene quasi continue di residui polari e acqua attraverso il braccio della membrana del complesso I. Queste catene collegano il sito di reazione dell’ubichinone a tre moduli separati «simili ad antiportatori» che spostano protoni attraverso la membrana. Nello stato chiuso post-reazione con ubichinolo legato, le connessioni appaiono in gran parte intatte ma punteggiate da piccole interruzioni dove l’acqua è probabilmente troppo mobile per essere vista, suggerendo percorsi flessibili piuttosto che tubi rigidi. Al contrario, uno stato alternativo aperto e deattivato—stabilizzato qui da una molecola di detergente incastrata nel canale—mostra un’ansa elicoidale che interrompe una connessione chiave, a sostegno dell’idea che questo stato regolatorio blocchi temporaneamente il trasferimento protonico su lunga distanza.
Cosa significa per la salute cellulare
Nel complesso, gli istantanee strutturali e le simulazioni supportano una sequenza in cui l’ubichinone si lega in una posa pre-reazione, accetta due elettroni e due protoni per diventare ubichinolo, e poi rimane in una posa post-reazione mentre i gruppi proteici vicini attendono di essere reprotonati dal lato matriciale della membrana. Il lavoro chiarisce come sottili cambiamenti nella posizione e nella carica di pochi atomi al sito attivo possano propagarsi attraverso lunghe vie protoniche per guidare il pompaggio, e come forme alternative, deattivate, possano interrompere questo flusso. Affinando la nostra visione del complesso I in azione, questi risultati forniscono un quadro per comprendere difetti ereditari, l’azione di farmaci e il danno mitocondriale che si verifica durante eventi come infarti e ictus.
Citazione: Chung, I., Pereira, C.S., Wright, J.J. et al. Post-catalysis structures of mitochondrial complex I with ubiquinol-10 bound in the active site. Nat Commun 17, 3506 (2026). https://doi.org/10.1038/s41467-026-70030-0
Parole chiave: complesso I mitocondriale, ubichinone-10, struttura cryo-EM, pompagGio di protoni, fosforilazione ossidativa